 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
African Crop Science Journal, Vol. 7. No. 4, pp. 321-326, 1999 RESISTANCE TO STRIGA ASIATICA (L.) KUNTZE IN SORGHUM: PARENT CHARACTERISATION AND COMBINING ABILITY ANALYSIS C.S. MUTENGWA, P. TONGOONA, S. MABASA1 and O. A.
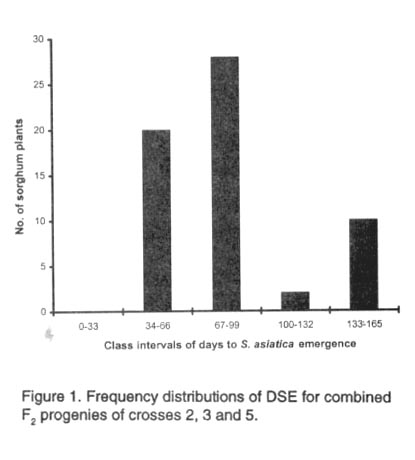
CHIVINGE Code Number: CS99021 ABSTRACT Pot experiments were conducted to assess the resistance of five sorghum [Sorghum bicolor (L.) Moench] cultivars namely, DC 75, SV-1, SAR 16, SAR 19 and SAR 29 to witchweed [Striga asiatica (L.) Kuntze] infestation. These genotypes were evaluated in two separate experiments, for resistance under S. asiatica infestation, and for yield without infestation. F2 progeny resulting from a four parent half diallel cross involving SV-1 and SAR cultivars were evaluated under S. asiatica infestation. The SAR and SV-1 cultivars were also screened for witchweed seed germination stimulant production using the agar gel technique. Cultivars SAR 19 and SAR 29 supported significantly lower S. asiatica counts than DC 75, SV-1 and SAR 16. They also delayed emergence of S. asiatica by more than 10 weeks relative to DC 75, SV-1 and SAR 16. Using agar assays, SAR 19 and SAR 29 had the least witchweed seed germination distances of 5.9 mm and 6.9 mm, respectively, compared to SAR 16 which had 15.4 mm and SV-1 which had 25.6 mm. Combining ability analysis for S. asiatica counts at maximum emergence revealed that additive genetic factors were preponderant for witchweed counts. Usually, SAR 16 is regarded as a resistant cultivar though its reaction to witchweed in these experiments revealed that it is susceptible. Cultivar SV-1 was susceptible, DC 75 was tolerant and SAR 19 and SAR 29 were resistant. Key Words: Diallel crosses, genetic effects, germination stimulant, recurrent selection, Witchweed, Sorghum bicolor RÉSUMÉ Des essais en pots ont été conduits pour évaluer la résistance de cinq cultivars de sorgho [Sorghum bicolor (L.) Moench] à savoir: DC 75, SV-1, SAR 16, SAR 19 et SAR 29 à l'infestation de l'herbe magicienne [Striga asiatica (L.) Kuntze]. Ces génotypes ont été évalués en deux essais séparés, pour la résistance à l'infestion de S. asiatica et pour le rendement sans infestation. La descendance F2 provenant des hybrides d'un demi diallèle de quatre parents comprenant des cultivars SV-1 et SAR ont été évalués sous l'infestion de S. asiatica. SV-1 et le cultivar SAR ont été aussi criblés pour la germination des semences de l'herbe magiciènne stimulant la production en utilisant la technique de gel agar. Les cultivars SAR 19 et SAR 29 ont supporté significativement un nombre faible de S. asiatica plus que DC 75, SV-1 et SAR 16. Ils ont aussi retardé l'émergence de S. asiatica plus de 10 semaines relativement à DC 75, SV-1 and SAR 16. Utilisant des essais agar, SAR 19 et SAR 29 ont eu la moindre germination des semences de la mauvaise herbe avec des distances de 5.9 mm et 6.9 mm respectivement, en comparaison avec SAR 16 qui avait 15.4 mm et SV-1 ayant eu 25.6 mm. L'analyse de l'aptitude de combinason pour les comptes de S. asiatica à l'émergence maximale a révélé que les facteurs génétiques additifs étaient préponderants pour les comptes de striga. SAR 16 est normallement considéré comme cultivar résistant bien que sa réaction au striga dans ces essais ont révélé qui'il était sensible. SV-1 était sensible, DC 75 tolérant alors que SAR 19 et SAR 29 étaient résistants. Mots Clés: Hybrides diallèle, effets génétiques, le stimulant de le germination, sélection recurrente, herbe magicienne, Sorghum bicolor INTRODUCTION One of the major biological constraints to increased sorghum productivity in the smallholder farming sector in Zimbabwe is attack by witchweeds, the most common of which is Striga asiatica (L.) Kuntze. Striga asiatica is a devastating obligate root parasite of cereal crops of the family Graminae (Stroud, 1993). Crop yield losses due to S. asiatica may be up to 100% depending on the extent of damage and level of infestation (Obilana and Ramaiah, 1992). Striga asiatica resistant cultivars offer an economically feasible and culturally sustainable technology for smallholder farmers since they do not require additional inputs (Mabasa, 1996). However, there are no high yielding S. asiatica resistant cultivars for resource-poor farmers in Zimbabwe. In sorghum, resistance results from one or a combination of several recognised mechanisms that influence the development of parasitism (Ejeta et al., 1993). The best characterised of these mechanisms is low stimulant production. This mechanism is inherited as a single, nuclear, recessive gene (Vogler et al.,1996). The production of germination stimulants by different genotypes is relatively simple to assay using the agar gel technique. In addition, it is noteworthy that low stimulant production is sufficient to confer field resistance to S. asiatica, independent of other resistance mechanisms (Hess et al., 1992). The concept of combining ability is useful to study and compare the performances of lines in hybrid combinations. It provides plant breeders with invaluable genetic information which enables them to choose the most appropriate selection criteria for desirable genotypes from both segregating and advanced breeding populations. The objectives of this work were to screen five sorghum cultivars for resistance to S. asiatica using both the pot culture and agar gel techniques and to determine the combining abilities of SV-1, SAR 16, SAR 19 and SAR 29 for S. asiatica counts. MATERIALS AND METHODS Pot experiments were conducted at Henderson Research Station (17º30,S and 31º E), 25 km from Harare. The sorghum cultivars comprised three S. asiatica resistant genotypes (SAR 16, SAR 19 and SAR 29) and two tolerant cultivars (SV-1 and DC 75) (Mabasa, 1996). The control was DC 75. A four-parent half diallel cross (Griffing, 1956) between the SAR cultivars and SV-1 was initiated during the 1995/96 rainy season. Since F1 generation seeds were not sufficient to conduct a comprehensive analysis, they were selfed during the 1996/97 rainy season to give six F2 populations which were used in this study. The following crosses were made; cross 1 (SAR 16 X SAR 19), cross 2 (SAR 16 X SAR 29), cross 3 (SAR 16 X SV-1), cross 4 (SAR 19 X SAR 29), cross 5 (SAR 19 X SV-1) and cross 6 (SAR 29 X SV-1). Plastic pots measuring 22.2 cm diameter and 22.3 cm deep were utilised for pot experimentation during the 1998/99 rainy season. Parental stocks were evaluated in two separate experiments, for resistance under S. asiatica infestation, and for yield without infestation. F2 progenies were only evaluated under S. asiatica infestation. About 0.02 g of S. asiatica seeds and 2 g of compound D fertiliser (7% N; 14% P205; 7% K20) were thoroughly mixed with the top 5 cm of soil prior to planting. Five sorghum seeds were subsequently planted per pot and thinning was done 2 weeks after crop emergence to leave one plant per pot. The experiments were laid out in randomised complete block designs with four replications per parent genotype and four replications for each F2 population. Statistical analyses were done on four pots per parent genotype and 20 pots for each F2 population. Sorghum grain yield was recorded from each pot. Witchweed components were days to first S. asiatica emergence (DSE) and S. asiatica counts per pot. Where S. asiatica did not emerge, days to physiological maturity of sorghum, estimated to be 160 days, was recorded as DSE. Witchweed counts at maximum emergence for each parent and F2 population were used for statistical analyses. Homogeneity of variance was tested for all the six populations to see if the distributions of DSE and S. asiatica counts per plant were uniform. Crosses 2, 3, and 5 had similar distributions for both parameters and they were thus combined and frequency distributions were plotted for 60 progenies for each parameter. Means were calculated for each class interval and standard error (SE) of means were only computed for classes with two or more cases (Panse and Sukhatme, 1985). The observed variation of witchweed counts among the F2 progenies was partitioned into general combining (GCA) and specific combining ability (SCA) effects following Griffing’s Model I, method 4 (Griffing, 1956). Agar assays were done following the methods described by Hess et al. (1992). Basically 100 ul preconditioned S. asiatica seeds were pipetted into petri dishes. Water agar (0.7%) was then poured over the S. asiatica seeds. Roots of germinating sorghum seeds were placed in the solidifying agar with the root tip pointing across the plate. The plates were incubated in the dark at 28oC for 72 hrs. Germinating S. asiatica seeds were easily visible through the bottom of the petri dish by viewing under a dissecting microscope. Maximum germination distance (MGD), that is, the distance between host root and the most distant germinated S. asiatica seed, and germination percentage, were used as indexes of resistance. The percentage of germinated witchweed seeds was determined by examining seeds in three 16 mm diameter microscope fields. The experiment was laid out in a completely randomised design with five replications per genotype. RESULTS Striga asiatica emergence. Days to S. asiatica emergence for the parent stocks ranged from 62 to 146 from sorghum planting. Cultivars SAR 19 and SAR 29 significantly (P<0.05) delayed the emergence of witchweeds by 10 weeks, compared to DC75, SAR 16 and SV-1 whose emergence was after 9 weeks from sorghum planting. Witchweeds began to emerge between 60.5 to 114.7 days for the six F2 populations and there were no significant differences (P<0.05) for DSE between plants. Most of the segregants from crosses 2, 3 and 5 had emergence occuring nine weeks from sorghum planting (Fig. 1) The mean (±SE) number of DSE for the four classes with S. asiatica emergence were: 34-66 (62.1±0.7; n=20); 67-99 (73.1±1.5; n=28); 100-132 (103.5; n=2); 133-165 (160±0.00; n=10); where n is the number of cases per class. All the progenies in class five did not have S. asiatica emergence such that DSE coincided with days to physiological maturity. The mean number of sorghum plants per class was 12±5.3. Striga asiatica counts. Cultivar SAR 29 reached maximum count after 101 days, SAR 19 after 126 days, DC 75 after 112 days and both SV-1 and SAR 16 after 119 days. Both SAR 19 and SAR 29 supported significantly lower numbers of S. asiatica plants than DC 75. SV-1 and SAR 16 had significantly (P < 0.01) lower Striga counts than DC 75, which had the highest mean count of 28.8 (Table 1). TABLE 1. Mean S. asiatica counts and days to Striga
emergence of five sorghum cultivars at Henderson Research Station, 1998/99 season
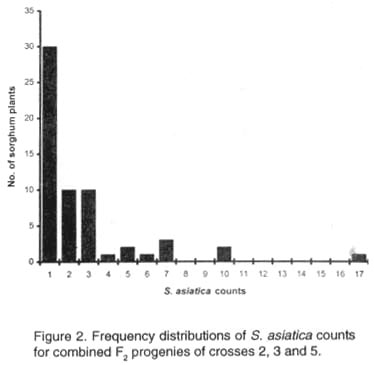
aData subjected to log10 (X + 1) transformation Most of the segregants from the combined population of crosses 2, 3 and 5 were resistant and this resulted in highly significant (P<0.001) skewing (3.09) and kurtosis (10.36) values (Fig. 2). The class interval was seven S. asiatica plants from zero for class one to 119 for class 17. The mean (±SE) witchweed counts for the nine classes with S. asiatica plants were: 0-7 (2.6±0.4; n=30); 8-14 (10.9±0.7; n=10); 15-21 (17.1±0.6; n=10); 22-28 (24; n=1); 29-35 (32.5±0.5; n=2); 36-42 (39±3.0; n=2); 43-49 (47.5±1.5; n=2); 64-70 (68.5±1.5; n=2); 113-119 (116; n=1); where n is the number of cases per class. The mean number of sorghum plants per class was 3.5±1.8. Combining ability analysis for S. asiatica counts. General combining ability components were highly significant (P<0.01) (Table 2). Thus, additive genetic factors were operative predominantly for witchweed counts. The GCA:SCA ratio was very large, 32.17 (Table 2), further revealing the greater influence of additive genes. Both SAR 16 and SV-1 had positive GCA effects while SAR 19 and SAR 29 had negative GCA effects (Table 3). The former cultivars consequently increased the number of S. asiatica plants in their F2 progeny. Conversely, SAR 19 and SAR 29 reduced the number of parasite counts in their progeny. TABLE 2. ANOVA table for S. asiatica counts at maximum
emergence according to Griffing’s model 1, method 4
*, **Significant at P<0.05 and P<0.01, respectively; ns = not significant TABLE 3. General combining ability effects and variances of S.
asiatica counts of four sorghum cultivars
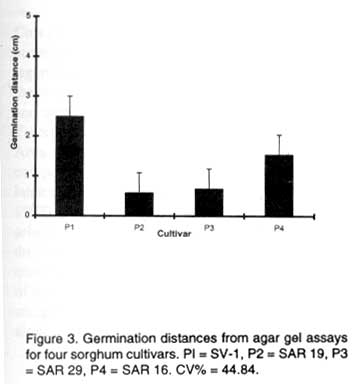
SE (gi-gj) = 0.110; LSD (0.01) = 0.257; LSD (0.05) = 0.188 Sorghum grain yield. Infestation with S. asiatica induced a highly significantly (P<0.01) yield loss for cultivars DC 75, SV-1 and SAR 16 (Table 4). Contrary to the original data, SAR 19 showed a significant (P<0.05) yield loss due to infestation because of the increased precision brought about by data transformation. Cultivar SAR 29 was the only cultivar that did not have a significant (P<0.01) yield reduction between infested and non-infested pots (Table 4). SV-1 had the greatest yield reduction of 81.7%, while SAR29 had the lowest yield reduction of 29.4% (Table 4). TABLE 4. Sorghum mean grain yields (g pot-1) for infested and uninfested cultivars
at Henderson Research Station, 1998/99 rainy season
aData subjected to square root transformation Agar gel assay. There was a high and positive correlation (r=0.88; P=0.001) between the percentage of S. asiatica seeds that germinated and the farthest distance at which they germinated after 72 hrs of incubation. The four sorghum cultivars differed significantly (P<0.01) for MSD. SV-1 was the highest germination stimulant producer with a distance of 2.56 cm, followed by SAR 16 with a distance of 1.54 cm (Fig. 3 ). Both SAR 19 and SAR 29 had witchweed seed germination distances which were less than 1 cm. DISCUSSION According to Hess et al. (1992), cultivars with a germination distance of less than 1 cm are resistant while those with more than 1 cm are susceptible. SV-1 and SAR 16 were thus susceptible. The magnitudes of yield loss for both SV-1 and SAR 16 under S. asiatica infestation also support this classification. These results are contrary to earlier reports in which SAR 16 has been reported to be resistant to S. asiatica (Obilana and Ramaiah, 1992; Mabasa, 1996). Cultivar DC 75 was tolerant, considering its high mean witchweed count and moderate yield reduction. Cultivars SAR 19 and SAR 29 supported the lowest S. asiatica counts, delayed emergence of S. asiatica by about 10 weeks and also had witchweed seed germination distances of less than 1 cm. These cultivars were therefore considered resistant. Their low grain yield potential should be improved through crossing them with high yielding cultivars or germplasm lines and selecting for both yield and resistance under witchweed infestation. The delay of witchweed emergence on SAR 29 may partly explain why this cultivar did not significantly lose its yield under Striga infestation (Gurney et al., 1999). Generally, most F2 progenies from the combined population of crosses 2, 3, and 5 delayed witchweed emergence and consequently had low parasites per plant. The highly significant GCA effects indicated that the performance of single-cross progeny can be adequately predicted on the basis of GCA. The most witchweed-resistant progeny can be produced by crossing cultivars with the least GCAs for witchweed counts. A recurrent selection programme or pedigree method with an emphasis on low witchweed numbers per plant could be adopted to evolve S. asiatica resistant cultivars. ACKNOWLEDGEMENTS Financial support for this work was provided by the Rockerfeller Foundation, and is highly appreciated. We are also indebted to T. Nyakandi and the Weed Research Team at Henderson Research Station for their technical assistance. The senior author also thanks D. Icishahayo for his guidance on data analysis and invaluable comments on the draft manuscript. REFERENCES
©1999, African Crop Science Society The following images related to this document are available:Photo images[cs99021a.jpg] [cs99021c.jpg] [cs99021b.jpg] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||

{kind=link}
{kind=link}
{kind=link}