 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
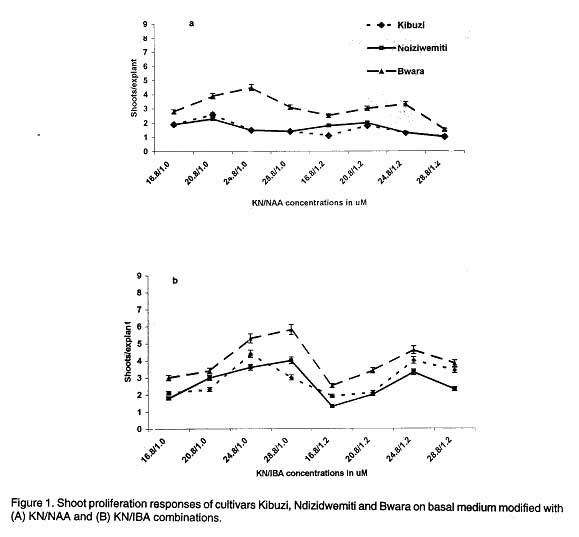
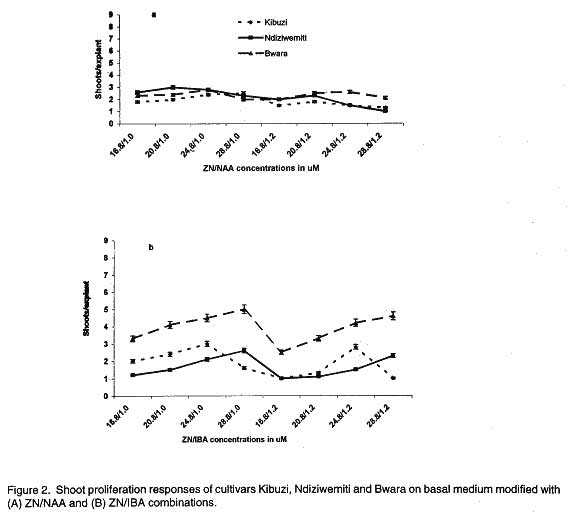
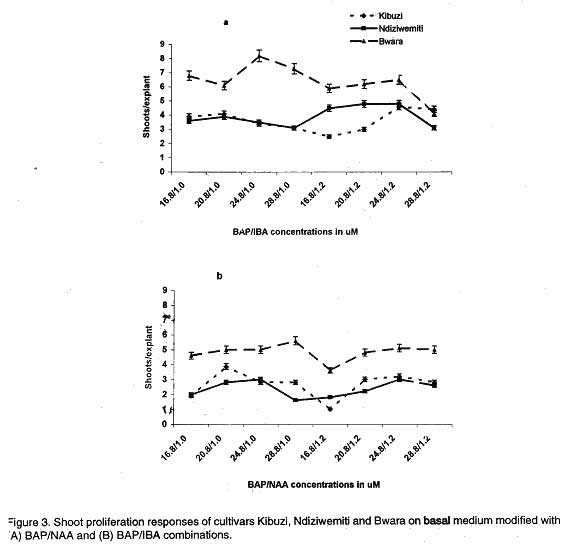
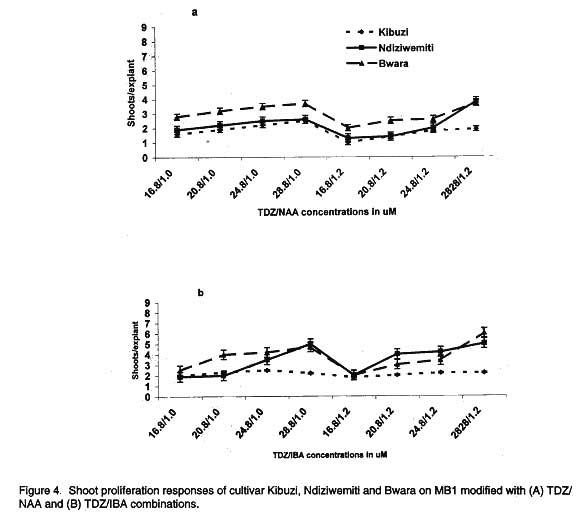
African Crop Science Journal, Vol. 7. No. 4, pp. 605- 611, 1999 Short Communication EFFECTS OF AUXIN/CYTOKININ COMBINATIONS ON SHOOT PROLIFERATION IN BANANA CULTIVARS G. Arinaitwe, P.R. Rubaihayo and M.J.S. Magambo Code Number: CS99052 ABSTRACT The effects of cytokinin/auxin interactions on in vitro shoot proliferation of bananas were investigated. Shoot-tips (3 mm), excised from in vitro-plantlets of Kibuzi (AAA-EA), Bwara (AAA-EA) and Ndiziwemiti (ABB) cultivars, were cultured on basal Murashige and Skoog modified medium (Talengera et al., 1994) to determine the cytokinin/auxin combination and concentration that gives high shoot proliferation in the three cultivars. The modified medium was supplemented variously by equimolar concentrations of Benzylaminopurine (BAP)/Indolebutyric acid (IBA): 16.8/1.0, 20.8/1.0, 24.8/1.0, 28.8/1.0, 16.8/1.2, 20.8/1.2, 24.8/1.2 and 28.8/1.2 µM. The above concentrations were also used in: Kinetin (KN)/ Napthalene acetic acid (NAA), Thidiazuron (TDZ)/NAA, TDZ/IBA, Zeatin (ZN)/NAA and ZN/IBA. ZN/NAA and TDZ/NAA supplemented medium suppressed shoot proliferation rates in the three cultivars at all concentrations while BAP/IBA combinations induced more shoot prolifration in cultivar Ndiziwemiti than was previously observed on 20.8 µM BAP modified MS medium. Cultivars reacted differently to different BAP/IBA concentrations: Ndiziwemiti had its best proliferation rate of 4.6 at 16.8/1.2 µM; Bwara, 8.5 at 24.8/1.0 µM; and Kibuzi, 4.5 at 24.8/1.2 µM. Other proliferation rates, for Ndiziwemiti, included 2.8 at 24.8 µM BAP/1.0 µM NAA and 4 at 28.8 µM / 1.0 µM IBA. In most cases higher proliferation rates were observed in cytokinin/auxin combinations in which a weak auxin (IBA) was included. Key Words: Auxins, cytokinins, Musa sp., shoot induction RÉSUMÉ Les effets de l’interaction entre la cytokinine et l’auxine sur la differantiation morphologique de la banane in vitro ont été étudiés. Les pousses de 3mm prelevées sur les plantules in vitro des cultivars Kibuzi (AAA-EA), Bwara (AAA-EA), et Ndizidwemiti (ABB) ont été cultivées sur le milieu basal de Murashige & Skoog modifié (Talengera et al., 1994) pour déterminer la combinaison et la concentration de cytokine/Auxine qui produit une grande prolifération des feuilles dans les trois cultivars. Le milieu modifié a été augmenté de concentrations variées d’equimolar de BAP/IBA (µM) suivantes: 16.8/1.0; 20.8/1.0, 24.8/1.0; 28.8/1.0; 16.8/1.2; 20.8/1.2; 24.8/1.2 et 28.8/1.2µM. Ces concentrations ont été aussi utilisées dans: Kinetine (KN)/Acide acétique de Napthalène (NAA), Thidiazuron (TDZ)/NAA, TDZ/IBA, Zeatine (ZN)/NAA et ZN/IBA. Les milieux supplémentés de ZN/NAA ont réduit le taux de prolifération dans les trois cultivars à toutes les concentrations. Les combinaison BAP/IBA a induit plus de prolifération de pousses du cultivar Ndiziwemiti que ce qu’ on a obtenu avec la concentration de 20.8 µM BAP du milieu Marashige-Skoog modifié. Les cultivars ont réagi differemment aux differentes concentrations de BAP/IBA: Ndiziwemiti a eu le plus meilleur taux de prolifération de 4.6 à 16.8/1.2µM Bwara, 8.5 à 24.8/1.0µM; et Kibuzi; 4.5 à 24.8/1.2µM. D’autres taux de prolifération pour Ndiziwemiti comprenaient 2.8 à 24.8µM BAP/1.0 µM NAA et 4 à 28.8 µM/1.0µM IBA. Générallement des taux élevés de proliferation ont été obtenus dans les combinaisons cytokinine/auxine dans lesquelles il était inclu de l’auxine (IBA) faible. Mots Clés: Auxines, cytokinines, Musa sp., induction des pousses INTRODUCTION Bananas and plantains (Musa spp.) are crops of significant economic importance in Uganda. They are a major source of food, income and employment to many Ugandans (Rubaihayo, 1991). Conventionally, they are propagated by suckers but this method is disadvantageous because suckers are usually infested and are a main source of weevils, nematodes and soil-borne pathogens (Sagi et al., 1998) and the low coefficient of multiplication (Vuylsteke, 1989). In vitro propagation through shoot-tip culture does overcome these problems. Different in vitro micropropagation protocols, using different adenine-based cytokinins, have been used in several Musa species of divergent genomic constitution and ploidy (Vuylsteke, 1989). Several workers have reported that cellular differentiation and organogenesis in tissue and organ culture are controlled by an interaction between auxin and cytokinin concentrations (Pierik, 1987; George and Sherrington, 1988; George, 1993; Razdan, 1993). George (1993) suggested that organogenesis in mono-cotyle-donous plants was promoted if cultured on media modified with cytokinin/weak auxin combi-nations. A balance between auxin and cytokinins is, therefore, required for adventitious and axillary shoot formation. Several growth regulant combinations have been used in non AAA-EA banana cultivars to induce multi-shoot growth, for example, 11.4 µM IAA/9.30 µM KN (Ma and Shii, 1972); 5.7 µM IAA/2.8 µM BAP (Cronauer and Krikorian, 1986); 9.9 µM IBA/23.3 µM KN (Bower, 1982); 11.4 µM IAA/22.2 µM BAP and 22.3 µM IBA/24.6 µM BAP (Dore Swamy et al., 1983). However, only 1 µM NAA/19.9 µM BAP combination was tested in a few AAA-EA banana cultivars (Talengera et al., 1994). This study reports on the influence of different auxin and cytokinin combinations on shoot proliferation of bananas using Talengera et al. (1994) medium which modified MS medium by supplementing it with 20.8 µM BAP as a standard. MATERIALS AND METHODS Shoot apices were excised from in vitro propagated cultures of three cultivars; Ndiziwemiti (ABB) which was reported to be recalcitrant to Talengera et al. (1994) modified medium with a proliferation rate of 1.5; Kibuzi (AAA-EA) with a mean proliferation rate of 3, and Bwara (AAA-EA) with a proliferation rate of 6 on the same medium. Three (3) mm shoot-tips were isolated aseptically and inoculated onto differently modified semi-solid nutrient media. The basal medium contained Murashige and Skoog (1962) macro- and micro-elements supplemented with 0.4 mgl-1 thiamin HCl, 0.5 mgl-1 nicotinic acid, 0.5 mgl-1 pyridoxine HCl, 100 mgl-1 myo-inositol and 2 mgl-1 ascorbic, and variously modified with equimolar concentrations of the different ratios of cytokinin/auxin combinations. Talengera et al. (1994) medium (MS modified medium) was used as a standard. The combinations used included: BAP/IBA (µM): 16.8/1.0, 20.8/1.0, 24.8/1.0, 28.8/1.0, 16.8/1.2, 20.8/1.2, 24.8/1.2 and 28.8/1.2. The above concentrations were also used in: KN/NAA, KN/IBA, ZN/NAA, ZN/IBA, BAP/NAA, TDZ/NAA and TDZ/IBA combinations. Cultures were incubated at 27± 2 oC under 16 hr photoperiod of 1770± 42 lux. Three weeks after inoculation, the shoot-tips had formed shoot clusters, which were sub-divided. Individual shoot-tips of comparable sizes were transferred onto fresh media. This procedure was repeated for three successive four-weeks-culture cycles. At the end of each culture cycle the number of shoots per explant (shoot proliferation rate) was counted and recorded. The data collected were analysed using MSTATC computer progra-mme and proliferation means separated by LSD method. RESULTS AND DISCUSSION After three weeks of culture on differently formulated media, the white in vitro explants turned green and the development of shoots began. Besides shoot and root proliferation, explants increased in size and callus was induced especially in high auxin/cyokinin combinations. More callus induction was observed in ZN/NAA (µM): 16.8/1.0 and 16.8/1.2. The cultivars’ shoot proliferation responses to KN/auxin combinations are presented in Figure 1. KN/NAA-supplemented medium (Fig. 1A) suppressed shoot proliferation rates of Kibuzi and Bwara compaired to their rates on standard medium (Talengera et al., 1994). Bwara appeared to be recovering its normal proliferation rate at 24.8/1.0 µM but was quickly suppressed at 28.8/1.0 and all the 1.2 µM NAA rates. All the KN/auxin combinations suppressed cultivar Kibuzi’s proliferation rates. Ndiziwemiti did not respond significantly to the different combinations of cytokinin and auxins compared to its performance on standard medium. Substituting NAA with IBA in the cytokinin-auxin combinations (Fig. 1B), changed the cultivars’ shoot proliferation reaction. Bwara significantly (P<0.05) proliferated better from 16.8/1.0 µM to 28.8/1.0 µM than on standard medium. Increasing IBA concentration to 1.2 µM in KN at 16.8/1.2 produced a lower proliferation rate but recovered with increasing KN rates. Increasing the KN concentration in KN/IBA combinations significantly (P<0.05) progressively increased Ndiziwemiti’s proliferation rate up to 24.8/1.2 µM after which it experienced a decline. Kibuzi showed a significant (P<0.05) increase in proliferation at 24.8/1.0 µM beyond which the rate reverted to 16.8/1.2 to 20.8/1.0 µM. Similar results were observed at 24.8/1.2 µM. Shoot proliferation responses of the three cultivars to ZN and auxin combinations and concentrations are presented in Figure 2. The proliferation of Kibuzi and Bwara at all concentrations of ZN/NAA (Fig. 2A) was significantly (P<0.05) suppressed compared to their proliferation on standard medium. However, Ndiziwemiti showed a significantly (P<0.05) higher rate on 20.8/1.0 to 28.8/1.0 µM and 20.8 /1.2 µM than on 20.8 µM BAP before declining. The ZN/IBA combination results are presented in Figure 2B. The shoot proliferation of all the cultivars were suppressed by most of the ZN/IBA combinations compared to the standard medium. Vuylsteke and De Langhe (1985) reported ZN as a weak cytokinin in Musa spp. shoot multiplication probably due to its susceptibility to cytokinin oxidases. Cultivar Bwara never showed any cytokinin/auxin combinations which performed to the level of the standard 20.8 µM BAP supplemented medium. In cultivar Kibuzi, only proliferation rates at 24.8/1.0 and 24.8/1.2 µM equaled the rate observed on the standard 20.8 µM BAP modified MS medium. Ndiziwemiti showed a significant (P<0.05) increase in proliferation rate at 28.8/1.0 and 28.8/1.2 µM concentrations. The BAP/IBA combinations and concentrations (Fig. 3A.) induced higher shoot proliferation rates compared to what was observed in other cytokinin/auxin combinations. Cultivar Bwara showed significant (P<0.05) increases in shoot proliferation at 24.8/1.0 µM and 28.8/1.0 µM concentrations compared to standard medium. IBA is a weak auxin and Sin Wan (1992) reported an increase in shoot proliferation as a result of strong cytokinin/weak auxin complementarity in Musa species. The other combinations did not cause significant reaction except combination of 28.8/1.2 µM which significantly (P<0.05) reduced the proliferation rate. All the BAP/NAA concentrations suppressed the proliferation rate of Bwara (Fig. 3B). Kibuzi did not show significant reaction from standard medium except for a significant (P<0.05) increase at 20.8/1.0 µM. Ndiziwemiti showed an increase in reaction to increasing BAP/IBA concentration from 16.8/1.0 to 24.8/1.0 and 20.8/1.2 to 28.8/1.2 µM. Complementing TDZ with NAA combinations at various concentrations (Fig. 4A.) suppressed Bwara and Kibuzi proliferation. Ndiziwemiti progressively increased its shoot proliferation rate on supplementation of 1.0 µM NAA but declined on supplementation of 1.2 µM NAA at 16.8 to 20.8 µM TDZ before recovering with increasing TDZ rates. Huetteman and Preece (1993) suggested that TDZ’s action was reduced by the addition of a strong auxin (NAA) and may explain the suppression of shoot proliferation at 1.2 µM NAA. Cultivar Ndiziwemiti showed a significant (P<0.05) increase in shoot proliferation rate although the rate was comparably lower than that observed when only TDZ was used (Arinaitwe et al., 1999). This type of reduction of proliferation rates as a result of some cytokinin/auxin supplementation in AAA-EA banana cultivars was also reported by Talengera et al. (1994). Complementation of the basal medium with TDZ/IBA (Fig. 4B) induced higher shoot proliferation reaction in Ndiziwemiti than with TDZ/NAA combinations. Ndiziwemiti showed proliferation rate increase with increasing TDZ concentration in the auxin combinations and significant (P<0.05) increases at 28.8/1.0 and 28.8/1.2 µM concentrations than the standard medium of Talengera et al. (1994). Increasing TDZ concentration in the TDZ/IBA combinations evoked higher shoot proliferation than what was observed on adenine-based cytokinin/auxin combinations. The shoot proliferation was manifested in appearance of numerous fleshy bulbous structures each of them producing several stunted tiny adventitious buds on their surfaces. High cytokinin activity of TDZ was earlier reported by Fellman et al. (1987) in woody plant species. Incorporation of a strong auxin in the media suppressed the shoot proliferation rates of the banana cultivars. On media modified with low cytokinin/auxin ratios, for example 16.8/1.0 and 16.8/1.2 ZN/NAA combinations, the East African Highland banana (AAA-EA) cultivars showed single shoot development and callus induction. This indicated increased level of auxin concentration within the explant tissues and hence increased apical dominance. Furthermore, the inoculation of AAA-EA banana cultivars on hormone free medium (Talengera et al., 1994) showed root development with limited shoot multiplication. Thus, the results of our study indicated the existence of high levels of endogenous auxins and, therefore, the added auxins in the media resulted in increased total auxin levels resulting in high apical dominance and hence suppression of shoot proliferation. The increased auxin level reduced the effect of the added cytokinin in promoting shoot proliferation since for in vitro proliferation, a high cytokinin/auxin ratio has to be maintained (George, 1993; Vuylsteke, 1989; Razdan, 1993). In most cases cytokinin/weak auxin (IBA) combinations induced higher shoot proliferation rates than cytokinin/strong auxin (NAA) combinations. George (1993) earlier reported increased shoot proliferation rates with cytokinin/weak auxin combination in other monocotyledonous plants. Comparing the shoot proliferation rates induced on media devoid of auxins (Arinaitwe, et al., 1999) and media with auxins, the AAA-EA cultivars can best be proliferated on auxin free media. The negative role of exogenous auxins application in shoot proliferation of bananas was also reported by Moteille and Foncelle (1989) in Poyo (AAA). Acknowledgement The work reported in this paper was supported by Forum grant from the Rockefeller Foundation. References
©1999, African Crop Science Society The following images related to this document are available:Photo images[cs99052c.jpg] [cs99052b.jpg] [cs99052d.jpg] [cs99052a.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}