 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
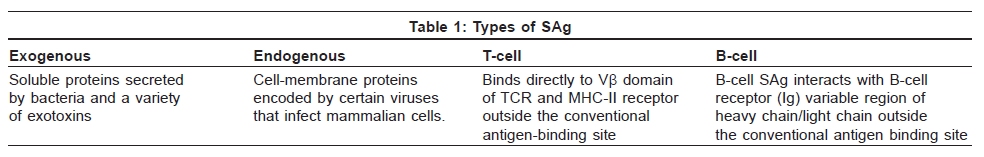
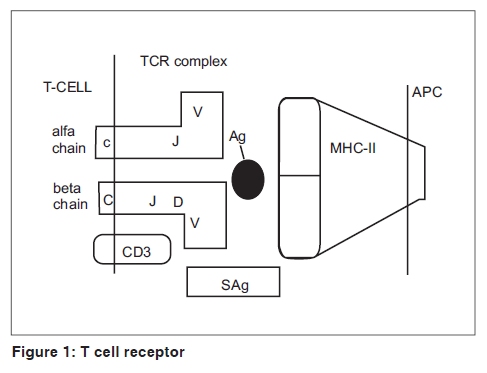
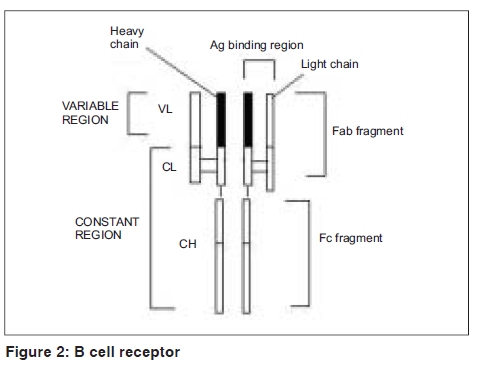
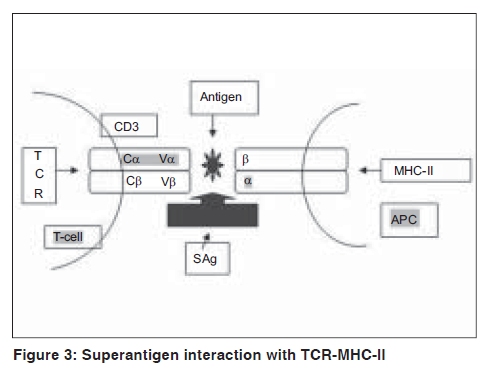
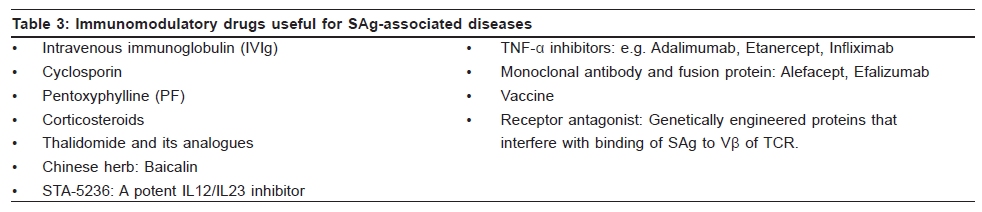
Indian Journal of Dermatology, Venereology and Leprology, Vol. 75, No. 5, September-October, 2009, pp. 540-544 Focus Superantigen Jignesh Vaishnani Department of Dermatology; SMIMER, Surat-395010; Gujarat, India Code Number: dv09185 PMID: 19736457 DOI: 10.4103/0378-6323.55423 Introduction Kappler et al . [1] describe a family of microbial proteins termed 'Superantigen' (SAg) that stimulates strong T-cell receptor (TCR) Vb restricted response. Superantigens are among the most potent T-cell mitogen known, with characteristic Vb signature. Previously SAg concept was limited to T cell only but recently the concept of B-cell SAg is growing. Besides classical SAg-mediated disease e.g. toxic shock syndrome (TSS), SAgs have also been proposed to contribute to the pathogenesis of several poorly understood acute and chronic inflammatory conditions including rheumatoid arthritis and psoriasis. Superantigens are not only powerful tools for the study of immunological phenomenon, but also its use is implicated in therapeutic intervention. Definition Superantigens are microbial proteins of 22-29 Daltons in size and are potent stimulators of the immune cells in a unconventional manner produced by bacteria, virus and mycoplasma. It has two domain folding comprising of the NH 2 terminal β barrel providing the binding region for the MHC-II receptor and α long COOH terminal a barrel providing the binding site for Vβ region of TCR [Table - 1]. T-Cell and B-Cell Receptors T-cell receptor (TCR) comprises of two peptide chains, either α/β or γ/δ, non-covalently associated with CD3γδε and ζ chains. [2],[3] Ninety percent of peripheral blood T cells have α/β peptide chain, while γ/δ chain is present on 4% (range 1-10%) of peripheral blood and lymph node lymphocytes and 1% of thymocytes. α Peptide chain contains three regions V (variable), J (junctional) and C (constant), while β peptide chain in addition has a fourth region D (diversity). Each TCR complex (α/β chain) constant region interacts with CD3γδε and ζ chains and consists of immunoglobulin like, connecting peptide, transmembrane and cytoplasmic domain [Figure - 1]. B-cell receptors B-cell receptors (BCRs) comprise of membrane-bound immunoglobulin (Ig) on the surface. The C region of Ig remains inserted in the membrane of B cell, while the V region acts as the antigen-binding site (Fab). For any given Ig molecule the V region differs from every other immunoglobulin (Ig). Sequence variability is found in three segments of V region, designated as hyper-variable regions e.g. V1, V2, V3, and identified in both heavy (VH) and light (VL). The most variable part of the region V is VH3 [Figure - 2]. Classical response In classical response, after antigen processing by antigen presenting cell (APC), an epitope from a protein antigen acts as a bridge between the HLA complex of APC and TCR. [2],[3] Only a small proportion of T cells become activated particularly after a co-stimulatory signal is produced by the APC. Response is highly regulated in order to limit harmful effects [Figure - 3]. Superantigen response T cell SAg binds directly to TCR and MHC-II receptor outside the conventional antigen-binding site, thus bypassing the restrictive feature of conventional antigen processing. [4],[5] Superantigen binds to Vβ domain of TCR, where Vβ refers to a variable region of b peptide. Different SAgs have specificity for one or limited sets of Vβ designation. SAg can stimulate all T cells bearing the particular Vβ designation, thus SAg can stimulate 20-30% of the total T lymphocytes in an individual [Figure - 3]. MHC-II positive cells are required for SAg-induced T-cell activation, but it is not MHC-II restrictive, and binding of MHC-II receptor determines the susceptibility of an individual to the particular SAg. Besides Vβ-specific T-cell activation, certain SAgs, e.g. SEH (Staphylococcal Enterotoxin H) induces Vα-specific T-cell activation. In case of MAM (mycoplasma arthritidis associated Superantigen), interaction is intermediate between SAg and conventional peptide antigen. B cell SAg [6] interacts with the variable region of heavy/light chain outside the conventional antigen-binding site, thus activating B cells in a VH selective manner. Most B-cell SAgs bind to the heavy chain from VH3 gene family. VH3 gene family is the largest of the seven human VH gene families and expressed by 30−60% of peripheral B cells [Table - 2]. Superantigen Interaction and its Effects As there is no definite disease model for SAg-mediated disease, many in vivo and in vitro studies demonstrate various effects of SAg. T-cell SAg Massive T-cell activation and release of cytokines, e.g. TNF-α, IL-2, IL-6, INF-γ in large amount, results in capillary leak and systemic shock. There is a biphasic response after SAg stimulation of T cell, with a T-cell derived initial peak of IL-2, TNF-α, followed by second peak from macrophage-derived cytokines.[7] Proliferation of Vß -specific T cell, but not an antigen restrictive. Deletion: [8] Initially Vβ-specific T-cells expansion followed by Vβ-specific clonal deletion of T cells. Anergy: [9] Hyporesponsive state of T cell to an antigen in the absence of appropriate co-stimulatory signal. T-cell dependent B-cell activation characterized by polyclonal IgM and IgG production, enhances humoral immunity via Ag-specific CD4 + T cells [10] Cytotoxicity: (1) Cyctotoxic T-cell mediated cytotoxicity against MHC class II positive cells, known as SAg-dependent cell-mediated cytotoxicity (SDCC).[11] (2) Activation-induced cell death (AICD).[12] (3) Superantigen-dependent autokilling. [13] Induction of autoimmune status: [14],[15] Although there are no direct evidences for this SAg has been proposed as one of the etiologies for autoimmune disease. Autoimmune state may result from indiscriminate Vb-specific expansion that amplifies the clone that manifests cross reactivity towards endogenous antigen and loss of self-tolerance. This may persist even after original SAg stimuli ceases. Three different mechanisms have been proposed for induction of autoimmune status (1) In presence of SAgs and multivalent autoantigen abnormal Th-B cell interactions lead to activation, proliferation and differentiation of B-cells and production of autoantibody. (2) T-cell independent and direct activation of B cell by SAgs. (3) Superantigens may activate resting T cells that recognize autoantigens and may remain in active state in the presence of autoantigen. Superantigens increase the expression of glucocorticoid receptor β and are associated with decreased corticosteroid response. [16] Other effects: [5],[10] Stimulates lymphocyte locomotion and neutrophilic recruitment to the site of infection, emesis and augmentation of endotoxin activity. Recruitment of T cells, B cells and APCs at the site of infection, and activation of B cells and APCs further augment the cytokine release. B-Cell Superantigen B-cell SAg binds to serum Igs and leads to the formation of large amount of immune complexes. Such immune complexes activate the complement pathway and inflict tissue injury. [6],[17] B-cell SAgs bind to the surface Igs on mast cells and basophils, resulting in the release of pro-inflammatory mediators. T-cell independent VH-specific B-cell activation and proliferation, followed by clonal deletion, and prolonged suppression of antibody production. Endogenous Superantigens Endogenous Superantigens (ESAgs) are cell membrane proteins encoded by certain viruses that infect mammalian cells. [18] In humans ESAg is encoded by env gene of human endogenous retrovirus (HERV), and all humans carry numerous copies of HERV in their genome. Exact significance of ESAg is not known in humans. Endogenous superantigen stimulates T cell in Vβ in a selective manner to support viral replication and plays a role in the pathogenesis of EB virus infections, HIV infection, CMV infection and IDDM (Insulin Dependent Diabetes Mellitus). Treatment Strategies for Superantigen -Mediated Disease As there is no definite disease model for SAg-mediated disease and lack of controlled trials about therapeutic intervention, many drugs are claimed to be effective with different immunological properties. Following treatment strategies are proposed for the diseases associated with SAg.
[Table - 4] gives superantigen toxins, and [Table - 5] diseases thought to be mediated by superantigen, and [Table - 6] factors affecting SAg-induced response. References
Copyright 2009 - Indian Journal of Dermatology, Venereology and Leprology The following images related to this document are available:Photo images[dv09185t5.jpg] [dv09185t4.jpg] [dv09185f1.jpg] [dv09185f2.jpg] [dv09185f3.jpg] [dv09185t2.jpg] [dv09185t6.jpg] [dv09185t3.jpg] [dv09185t1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}