 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Electronic Journal of Biotechnology, Vol. 3, No. 2, August, 2000 SHORT COMMUNICATION Protocol for regeneration in vitro of Arachis hypogaea L. María Florencia Cucco1 and Alberto Daniel Rossi Jaume*2 1Departamento
de Microbiología e Inmunología, Universidad Nacional de Río
Cuarto, Ruta 8 Km. 601 - 5800 Río Cuarto, Argentina. Tel: 0358-4676200
E-mail: florcucco@infovia.com.ar
Financial Support: Secretary of Science and Technology, Universidad Nacional de Río Cuarto. Received
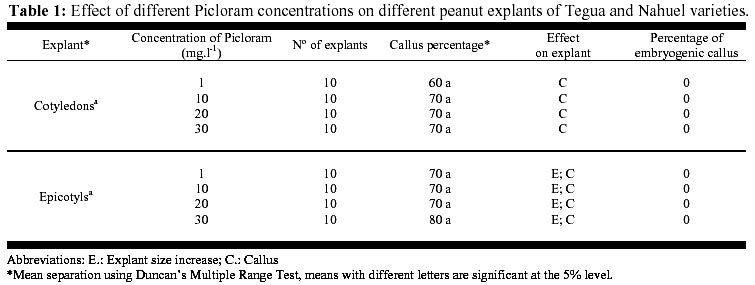
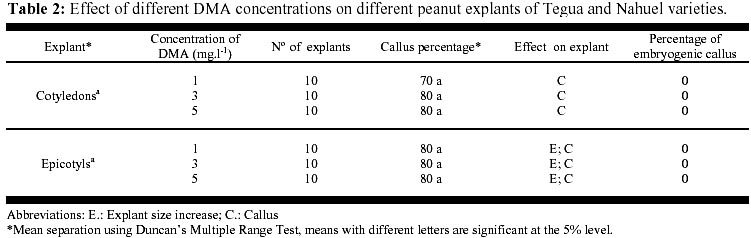
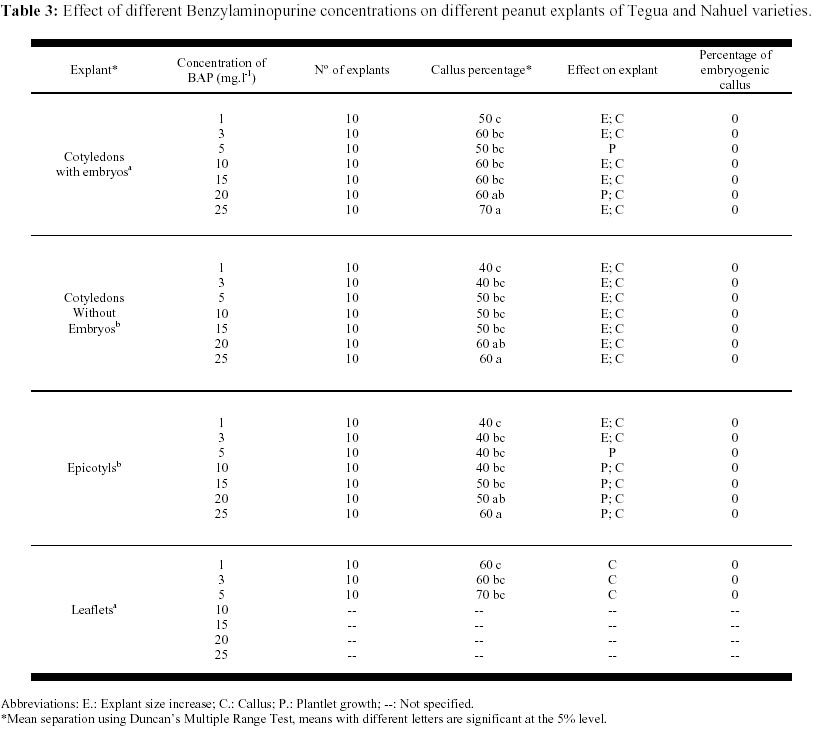
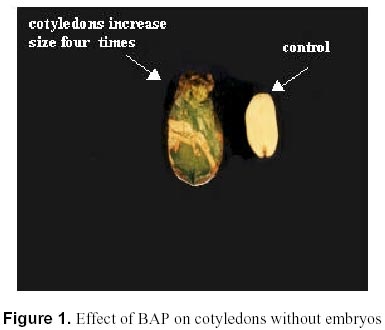
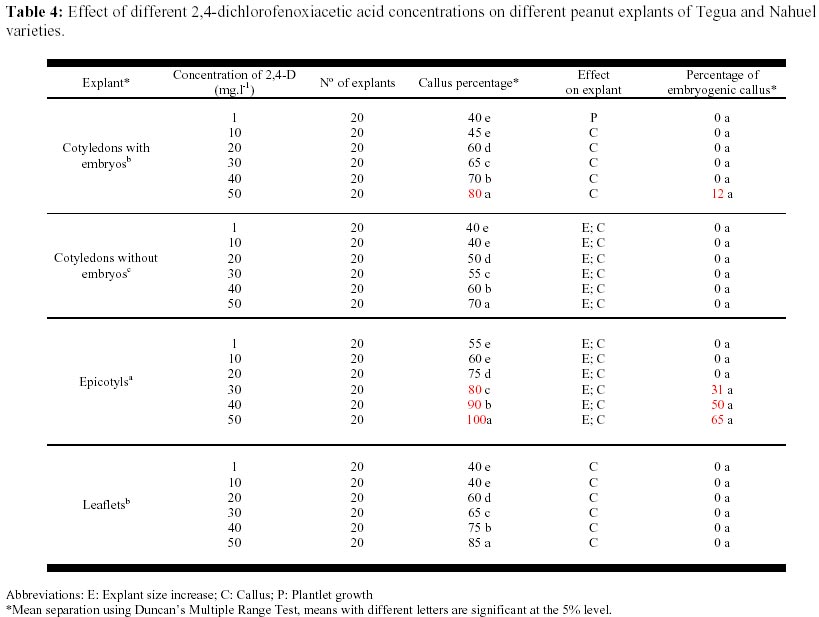
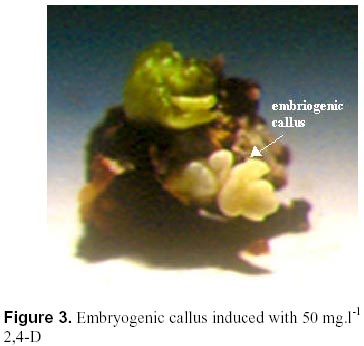
May 19, 2000 Code Number: ej00015 Abstract The regeneration of peanut Runner varieties grown in the South of the province of Cordoba, Argentina, was studied to count on a regeneration protocol, which is essential to perform genetic transformation of the plant. The induction of somatic embryos was evaluated in different peanut explants cultivated in Cordoba: cotyledons with and without embryos, epycotils and leaflets, of the Tegua, Nahuel and Florman INTA varieties. The induction medium consisted of MS salts with 30 g.l-1 sucrose, supplemented with the following hormones and herbicides: BAP, Picloram, DMA and 2,4-D. The highest percentage of embryogenic callus was produced cultivating epicotyls with 50 mg.l-1 2,4-D. The embryos obtained produced plantlets which, once acclimatized, reached the reproductive stage. Keywords: Growth regulators, Herbicides, Somatic embryogenesis. Abbreviations: MS: Murashige and Skoog medium (1962); 2,4-D: 2,4-dichlorophenoxyacetic acid; BAP: Benzylaminopurine; NAA: Naphthaleneacetic acid; Picloram [Picloram acid (4-amino-3, 5, 6 trichloropicolinic acid)], DMA (Dimetilamine salts of 3,6-dichloro-2-methoxybenzoic acid); INTA: Instituto Nacional de Tecnología Agropecuaria. Article The peanut varieties mainly grown in the south of the province of Cordoba (33º 0.6’ S and 64º 20’ W, 435 m.s.n.m), Argentina are the Runner varieties. One of the main problems of the peanut plant are its diseases, which are produced by the convergence of a susceptible cultivar, a virulent pathogen (fungus, bacteria, virus) and a favorable environment determined by the climate, soil and production system developed by humans. Among the main diseases that affect foliage, early and/or late smallpox is the most important. The sanitary control of peanut in our country has been organized in relation to this disease due to the great losses that it produces (Soave, 1997). Root diseases caused by soil fungi, such as smudges, withering, fruit rotting, have gradually increased their incidence and severity since the mid-eighties and constitute an important sanitary problem. With the development of different methods of genetic transformation, it has been possible to introduce genes with characteristics beneficial for the plant. Recombinant DNA technologies have greatly improved the quality of harvests by inserting genes that produce resistance to diseases caused by fungi or viruses in tomato, potato, and strawberry; resistance to herbicides in soybean, maize and oilseed rape; resistance to insects in cotton, potato and maize; genes that produce delay in fruit senescence in tomato; and genes that improve oil quality in oilseed rape (Delucchi, 1997). One of the conditions for plant transformation is to count on a good regeneration protocol. Regeneration through tissue cultivation can be mainly achieved through somatic organogenesis and embryogenesis. Numerous protocols of in vitro peanut regeneration have been reported in the last decade. Many protocols describe somatic embryogenesis using a great variety of explants such as leaflets (Baker and Wetzstein, 1992), immature cotyledons (Durham and Parrott, 1992; Ozias-Akins et al. 1992; Baker and Wetzstein, 1995); axes of immature embryos (Hazra et al. 1989; Ozias-Akins et al. 1992); hypocotyls (McKently, 1990; Li et al. 1994; Venkatachalam et al. 1997) and epicotyls (Rani and Reddy, 1997). Somatic embryogenesis has been described for more than one hundred species (Williams and Maheswaran, 1986; Tautorus et al. 1991) and in most of them the presence of auxins in the culture medium, especially the synthetic auxin 2,4-D, is the factor that determines the induction of embryo formation. This paper describes the regeneration of the Runner Florman, Tegua and Nahuel varieties using different explants, hormones and herbicides. Materials and methods Mature, dry seeds of high purity variety of Tegua, Nahuel and Florman INTA cultivars, obtained from "El Carmen" nursery, were used in the experience. Explants, cotyledons with and without embryos, epicotyls and leaflets were disinfected in 10% commercial bleach (55 g.l-1 active chlorum) for 10 min. with constant agitation and then rinsed three times with sterile water. After removal of the tegument, the same disinfection process was repeated. The embryos were extracted, the epicotyls (axis with leaflet and bud) were cut, and they were placed again in the same disinfecting solution for 5 min. with constant agitation and rinsed three times with sterile water. The cotyledons with and without embryos were disinfected following the same procedure. Leaflets were obtained from sterile plantlets grown in Magenta boxes. The leaflets used were between 10 and 15 mm long, and they were cut keeping the petiole. The leaflets were disinfected in 7% commercial bleach for 5 min. and then rinsed three times with sterile water. To study the effect of the different growth regulators on the induction of somatic embryos, the different explants were put in test tubes containing 15 ml of the following culture media: MS salts and vitamins, 30 gr.l-1 sucrose, 7 gr.1-1 agar (pH: 5.8) supplemented with hormones and herbicides: BAP (1, 3, 5, 10, 15, 20, 25 mg.1-1), Picloram (1, 10, 20, 30 mg.1-1), DMA (1, 3, 5 mg.1-1) and 2,4-D (1, 10, 20, 30, 40, 50 mg.1-1). The media were autoclaved at 121º C for 15 min. The tissues were stored in a phytotrom at 28º C with permanent photosynthetic active light at 38.6 µmol.m2.s-1. They were transferred to fresh induction media and callus formation was evaluated. Callus were placed in embryo growth and differentiation medium consisting of MS salts and vitamins supplemented with 30 gr.l-1 maltose, and incubated under the same conditions described above. The callus were transferred to fresh media every 30 days and the embryo formation was evaluated. The resulting somatic embryos were removed from the callus and cultivated in an MS medium supplemented with 60 gr.l-1 maltose, a medium which is adequate for the physiological maturation of the embryos. Finally, the mature embryos were transferred to an ½ MS medium, with 15 gr.l-1 sucrose and 0.1 mg.l-1 NAA for growth and rooting. Once the plantlets reached around 10 cm and had developed the necessary number of roots, they were planted in flasks with sterile soil and sealed with a plastic film, which was gradually removed to contribute to acclimatization. The plants were planted in flower pots, where they developed normally. Data were analyzed using the GLM procedure of the Statistical Analysis System (SAS) SAS Institute, (1990) and Ramos and Abbiati (1989). Mean separation was performed using Duncan's Multiple Range Test. Separation for mean number of embryos per explant or per embryogenic explant were performed after transforming embryo count data with square root of (emb. count + 0,5). Results The explants treated with Picloram herbicide, produced a high percentage of callus and epicotyl size increase (Table 1). There were no statistically significant differences in the production of callus among the different types of explants and the concentrations used. The callus placed in the embryo growth and differentiation medium did not produce embryos. The results obtained with DMA agrochemical were similar to those obtained with Picloram herbicide. In 1, 3 and 5 mg.l-1 concentrations, the treatment with DMA produced callus for both types of explants and epicotyl size increase. There were no statistically significant differences between the different explants (Table 2). The callus placed in the embryo growth and differentiation medium did not produce embryos. The different explants treated with BAP produced callus and epicotyl and cotyledon size increase for all the concentrations used. Cotyledons increased weight and volume up to four times (Table 3, Figure 1). The cotyledons with embryos and epicotyls, treated with BAP concentrations above 5 mg.l-1 induced plantlet growth breaking the apical dominance, which resulted in the growth of the axilar buds of the leaves (Figure 2). The cotyledons with embryos and leaflets produced more callus than cotyledons without embryos and epicotyls. The high concentrations of BAP produced a significant higher percentage of callus than the lower concentrations. The callus from different explants treated with BAP placed in an embryo growth and differentiation medium did not register embryo production (Table 3). When the explants were treated with different 2,4-D concentrations, they produced a high percentage of callus for both types of explants and size increase cotyledons with and without cotyledons and epicotyls (Table 4). Only those callus from cotyledons with embryos and epicotyls produced embryos in the growth medium and embryo differentiation (Figure 3). The explants of epicotyls produced a significantly higher number of embryogenic callus. In Table 5 it can be observed that the epicotyls of the Florman INTA variety, treated with 50 mg.l-1 2,4-D, produced embryos in percentages similar to those obtained for the varieties Tegua and Nahuel. Discussion and conclusion The analysis of the results obtained with different regulators and herbicides used for different explants of the Runner variety showed that the herbicides Picloram and DMA, both with auxinic effect, produced callus but these did not produce embryos. We suggest that the additives included in their formula may have inhibited embryogenesis. There were significant differences in the formation of callus in explants treated with BAP. The cotyledons with embryos and the leaflets produced the highest percentages of callus, but no hormone concentration applied to different explants produced embryogenic or organogenic callus. We have not come across studies reporting the induction of the formation of embryos in peanut by cytokinins. These are considered essential for embryo maturation once these are formed (Fujimura and Komamine, 1980). In the treatment with 2,4-D, the explants that produced the highest percentage of embryogenic callus were the epicotyls, followed by cotyledons with embryos, probably due to the fact that they carried embryos. This result agrees with the results reported by Rani and Reddy (1997), who obtained higher percentages of embryogenic callus cultivating cotyledons. The optimum concentration for the production of callus was 40 and 50 mg l-1, given that as the 2,4-D concentration decreases, the percentage of callogenesis also decreases. This result is in agreement with the findings of Baker and Wetzstein in 1994. The only hormonal treatment leading to somatic embryo formation was 2,4-D. This result is similar to that reported by Parrott (1998), who observed that the same Runner varieties are embryogenic but not organogenic. Embryogenesis is induced by compounds such as 2,4-D and other compounds of auxinic effect such as picloram (McKently, 1990). The epicotyls of the Florman INTA variety in a 50 mg l-1 2,4-D produced a percentage of embryogenic callus similar to those obtained with the Tegua and Nahuel Runner varieties. This may be due to the fact that the three varieties tested were produced from the same variety, Florrunner, from The United States, and was introduced in Argentina in 1977 (Soave, 1997). We conclude that 2,4-D is a compound that can produce somatic embryos from epicotyls of madure seed of the varieties Tegua, Nahuel and Florman INTA and generate plants that develop normally (Figure 4, Figure 5). Acknowledgements We thank Mercedes Ibañez, Licenciada en Biología, for assistance with the statistic analysis. References
Supported by UNESCO / MIRCEN network © 2000 by Universidad Católica de Valparaíso -- Chile The following images related to this document are available:Photo images[ej00015t5.jpg] [ej00015t1.jpg] [ej00015t4.jpg] [ej00015f5.jpg] [ej00015f3.jpg] [ej00015f2.jpg] [ej00015t2.jpg] [ej00015t3.jpg] [ej00015f1.jpg] [ej00015f4.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}