 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Electronic Journal of Biotechnology, Vol. 3, No. 2, August, 2000 SHORT COMMUNICATION Mitotic aberrations in coffee (Coffea arabica cv. 'Catimor') leaf explants and their derived embryogenic calli Andrea Menéndez-Yuffá *1, Rafael Fernandez Da Silva2, Liliana Rios3 and Nereida Xena de Enrech4 1Facultad
de Ciencias, Instituto de Biología Experimental, Laboratorio de Biotecnología
Vegetal, Universidad Central de Venezuela, Apartado 47114, Los Chaguaramos, Caracas
1041, Venezuela, Fax 7535897, E-mail: amenendez@cantv.net Received
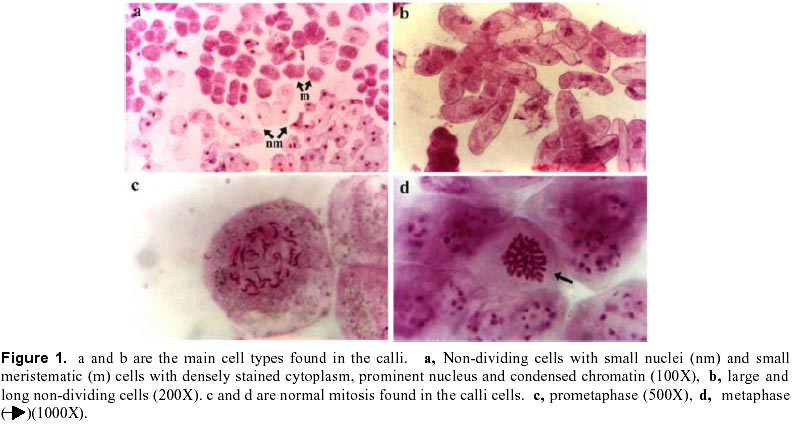
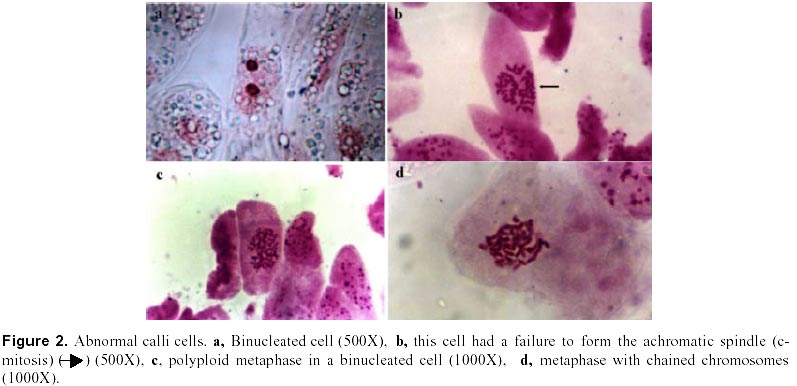
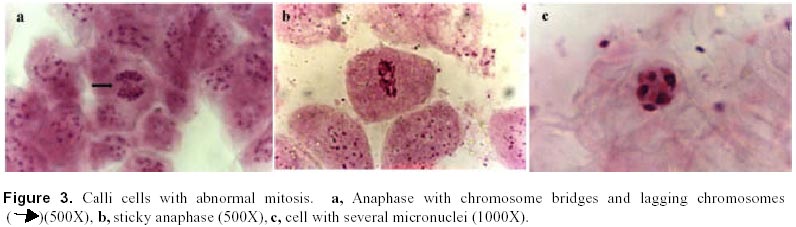
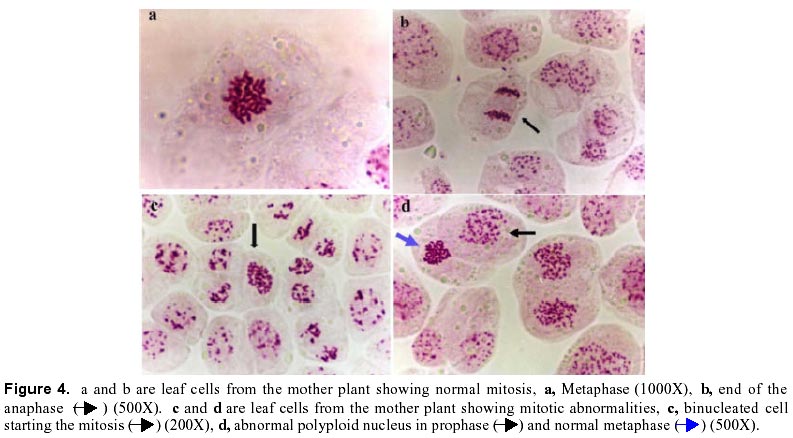
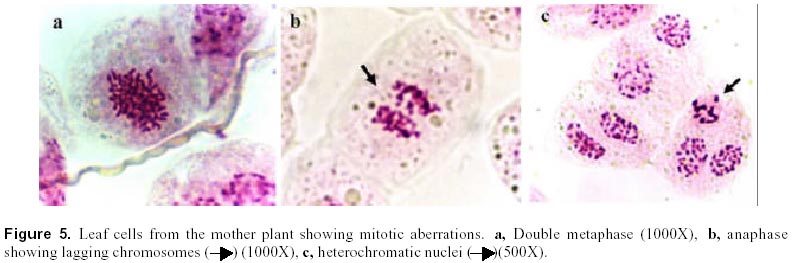
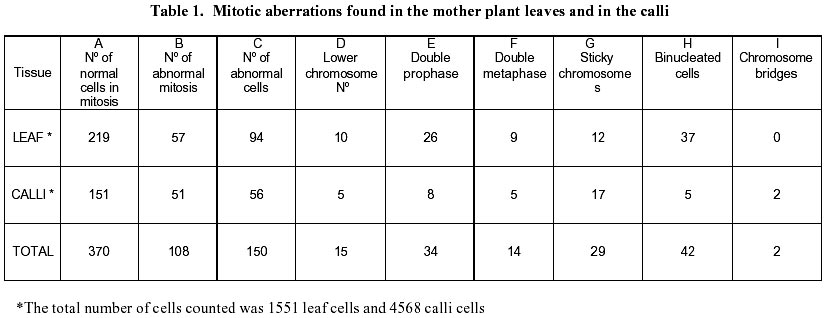
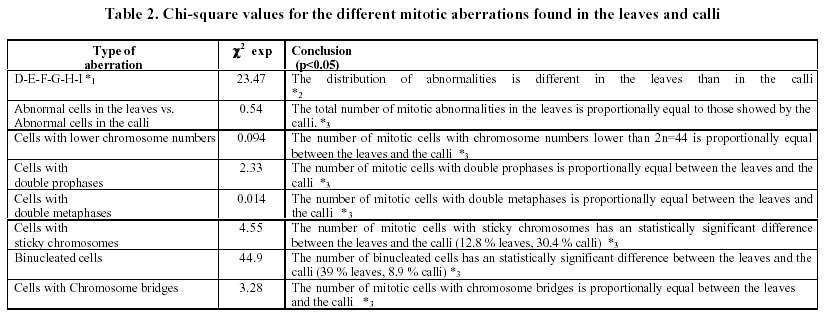
December 7, 1999 Code Number: ej00016 Abstract Dividing cells of leaves used as sources of explants from coffee plants (Coffea arabica cv. 'Catimor') and those of their derived calli were analyzed for mitotic aberrations. The studied tissues were prepared by squashing and stained with carbolfuchsin. A total of 1551 leaf and 4568 callus cells were surveyed. The majority (79 %) of leaf and calli (75 %) cells showed normal mitosis, however, cells with mitotic abnormalities were also found in both tissues. These included: polyploids, aneuploids, sticky chromosomes, double prophases and lagging chromosomes. Additionally, interphase cells with micronuclei or binucleated were also observed. The frequencies of these abnormalities were statistically different in calli and leaves. Calli showed a few other abnormalities such as c-mitosis, chained chromosomes, multipolar metaphases and chromosome bridges. Therefore, we conclude that these pre-existing abnormalities originate by errors in the process of normal mitosis in both leaves and in calli, and are therefore not caused by tissue culture conditions. Keywords: Coffee, Cytogenetics, Mitotic aberrations, Plant tissue culture, Somatic embryogenesis. Article Since the first publication of Staritsky in 1970, extensive work has been done on the induction of somatic embryogenesis in coffee, but very little is known about the genetic stability of the regenerated plants. In addition, the occurrence of potentially mutagenic mitotic aberrations in plant tissue cultures is well known (Evans et al., 1984; Sree Ramulu et al., 1985; Karp and Bright, 1985; Lee and Phillips, 1988). This study is part of an investigation on the genetic stability of coffee somatic embryos and the plantlets derived from them. Since part of the variation in plant tissue cultures, particularly in callus cultures, has been attributed to mitotic aberrations (Larkin and Scowcroft, 1981), these aberrations were analyzed in calli and leaves from coffee plants. The objective was to assess their influence in the stability of the regenerated plants and also to determine if the mitotic aberrations found in the calli were caused by tissue culture conditions. Materials and Methods Tissue culture. Somatic embryogenesis was induced according to García and Menéndez (1987). For callus induction, leaf sections of coffee (Coffea arabica cv. 'Catimor') were cultured for four months in medium containing half strength Murashige and Skoog (1962) salts, 30 g/l sucrose, 10 mg/l thiamine, 100 mg/l myo-inositol, 35 mg/l cysteine, 1 mg/l 2,4-dichlorophenoxyacetic acid, 8 mg/l 6-bencyladenine and 8 g/l agar. The resulting calli were transferred to a medium of similar composition but with 0.8 mg/l naphthalene acetic acid as a sole plant growth regulator. Chromosome squashes. Chromosome squashes were prepared from immature leaves of adult plants used for tissue culture and 3 months old embryogenic calli. The plant tissues were fixed for at least 12-24 hours in Carnoy (1 part of acetic acid: 3 parts of ethanol) followed by hydrolysis in 1 N HCl for 30-60 minutes and then placed on slides and stained with carbolfuchsin during 30-60 minutes. The tissue was squashed under a coverslip in a drop of 50 % glycerine. Statistical analysis. A total of 1551 leaf cells and 4568 calli cells were surveyed, from these, the interphase cells of normal appearance were discounted. The remaining cells were classified as normal mitotic cells, abnormal mitotic cells or binucleated cells. The cells in the second group were categorised by mitotic abnormality. A data matrix was constructed with this information, and tested for goodness-of-fit using the chi-square test, comparing the proportion of mitotic aberrations present in the leaves with the proportion of those found in the calli. Results and Discussion The embryogenic calli showed several types of cells (Fig. 1a and 1b) including non-dividing cells with small rounded nuclei (Fig. 1a), meristematic cells (Fig. 1a), which are small with densely stained cytoplasm and a prominent nucleus with condensed chromatin, and parenchyma cells, which are large and elongated (Fig. 1b). All these types of cells are characteristic of calli and have been previously described by Yeoman and Street (1973). Considering the numerous reports of mitotic aberrations in calli (Mythili et al. 1995; Ezura and Oozawa, 1994; Fluminhan and Kameya, 1996; Joachimiak et al., 1995; Lee and Phillips, 1988), especially in tissues incubated in vitro during long periods of time, we were interested in the evaluation of this tissue. The chromosome squashes of calli showed the majority of cells (75 %) undergoing normal mitosis (Fig. 1c and 1d) and a 25 % of the cells displayed abnormal mitosis, including changes in chromosome numbers such as aneuploids with chromosome numbers lower than 2n = 44, and polyploids (Fig. 2c), cells with chromosomal aberrations such as chromosome chains (Fig. 2d), bridges (Fig. 3a), lagging (Fig. 3a), and sticky chromosomes (Fig. 3b). Cells with double prophases, multipolar metaphases, c-mitosis (Fig. 2b), micronuclei and binucleated cells (Fig. 2a, 2c and 3c), were also observed. Some of these mitotic aberrations have also been reported in embryogenic calli of maize (Fluminhan and Kameya, 1996), and potato (Sree Ramulu et al., 1985), in cell suspensions of Daucus (Bayliss, 1975) and in cultured cells of Rauwolfia (Kunakh, 1996). To establish whether the mitotic aberrations observed in the calli were caused by the tissue culture conditions or whether they were previously present in the explant, leaf tissues from the donor plants were cytologically examined. The majority (79 %) of leaf cells showed normal mitosis (Fig. 4a, 4b) but the rest also displayed most the same type of mitotic aberrations observed in the calli (Fig. 4c, 4d, 5a, 5b and 5c). The frequency of each aberration is shown in Table 1. The total number of cells displaying mitotic aberrations in calli was not significantly different from those in leaves, but their distribution was different (Table 2). The frequency of double prophases, double metaphases, chromosome bridges and cells with lower chromosome number was statistically the same for both tissues, whereas binucleated cells were more frequent in leaves and the sticky chromosomes were more frequent in calli. In conclusion, coffee leaves have a number of evident mitotic aberrations that are also present in calli derived from them. The type and frequencies of such mitotic aberrations were similar in both tissues. Therefore, we conclude that these are pre-existing abnormalities, which originate by errors in the process of normal mitosis in both leaves and in calli, and were not caused by tissue culture conditions. The high frequency of mitotic aberrations in these tissues is in contrast with the stable chromosome number observed in somatic embryos and plantlets (Xena de Enrech et al., 1996). This finding suggests that most of the abnormal cells are incapable of regeneration and that there is autoselection of normal cells, which are capable of differentiating into somatic embryos. Considering that there are numerous genetic changes that cannot be seen through cytogenetical observations, it is advisable to make further analyses of the somatic embryos and the resulting plantlets by other methods, such as DNA sequence polymorphism and agronomic evaluations. These will allow to confirming the genetic stability of these plants. Acknowledgements The authors wish to thank Lic. Luis Hermoso by his work in the maintenance of plant tissue cultures, to Dr. Ana Herrera and Dr. Carlos F. Quiros for the revision of the manuscript, also to the "Consejo de Desarrollo Científico y Humanístico" (Project Nº 03.33.4049.97) and CONICIT (project Nº 2078) for the financial support. References
Supported by UNESCO / MIRCEN network © 2000 by Universidad Católica de Valparaíso -- Chile The following images related to this document are available:Photo images[ej00016f4.jpg] [ej00016t2.jpg] [ej00016f3.jpg] [ej00016f5.jpg] [ej00016f2.jpg] [ej00016f1.jpg] [ej00016t1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}