 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Electronic Journal of Biotechnology, Vol. 4, No. 1, April, 2001 Purification of a functional competitive antagonist for calcitonin gene related peptide action from sardine hydrolysates Marthe Rousseau1, Irineu Batista2, Yves Le Gal3 and Martine Fouchereau-Peron*4 1Station
de Biologie Marine, Muséum National d'Histoire Naturelle, CNRS FRE 2125 BP 225
29182, Concarneau Cedex, France E-mail: rousseam@gmx.net Financial support: This work was supported by a grant (FAIR CT 97-3097) from the European Community. Received
November 17, 2000 Code Number: ej01004 Abstract Calcitonin gene related peptide (CGRP) related molecules were purified from sardine hydrolysates prepared using 0.1% alcalase and two hours of hydrolysis. Gel exclusion chromatography and HPLC performed purification of these molecules. The purified molecules were characterised using specific CGRP radioimmunoassays and radioreceptoraasays. From 22 mg of crude extract, we obtained 14 µg of CGRP related molecules, the molecular weight determined by mass spectrophotometry was 6000 daltons. The biological activity of these molecules was analysed using the ability of CGRP to stimulate the adenylate cyclase activity in rat liver membranes. The purified molecules induced an inhibition of the CGRP stimulated adenylate cyclase activity, this effect was specific as no such effect was observed on the glucagon stimulated adenylate cyclase activity measured in the same rat liver membrane preparation. These results suggest that the purified molecules may act as antagonists for peptides that bind to CGRP receptors in rat liver membranes. These new antagonists may be of particular importance in various aspects of CGRP action in vertebrates. Keywords: biological activity, cyclic AMP, peptone, radioimmunoassay, radioreceptorassay. Article Many synthetic compounds as well as almost all organisms including animal, plants and microorganisms have been screened for bioactive substances as starting materials for biomedicines. Although numerous substances have been isolated, there are probably many others, especially degradation products of proteins that have not been identified. Through a control of process parameters such as pH, time and enzyme-substrate ratio, it is possible to produce hydrolysates whose components may have retained various functional properties attached to the native molecules (Adler-Nissen, 1982). Previous experiments performed using a large range of hydrolysates from various sources demonstrate the presence of CGRP immunorelated molecules in various fish hydrolysates (Fouchereau-Peron et al. 1999). The calcitonin gene related peptide (CGRP), a 37 aminoacid neuropeptide, is derived from the same gene as calcitonin by a mechanism of alternative splicing (Amara et al. 1982). It is predominantly synthesised in neural tissue and is mainly involved in the control of vasodilatation, with inotropic and chronotropic effects on the heart (Franco-Cereceda et al. 1987) but is also implicated in the regulation of gastric acid secretion (Hughes et al. 1984; Lenz et al. 1984). This peptide can also inhibit the proliferative response of T lymphocytes to mitogens (Umeda and Arizawa, 1989) and macrophage activation (Nong et al. 1989). In addition, at high doses, CGRP induces the same effects as calcitonin, that is hypocalcemia and hypophosphatemia (Roos et al. 1986). In non mammalian vertebrates, this peptide is mainly found in gills and intestine (Fouchereau-Peron et al. 1990b) and is involved in the control of hydromineral metabolism by its specific action on gill membranes (Fouchereau-Peron et al. 1990a; Arlot-Bonnemains et al. 1991). The sequence similarity between calcitonin and CGRP suggests that both peptides may support identical biological effects mainly in the control of homeostasis and reproduction (Lopez et al. 1976; Bjornsson et al. 1986). Among the various tested hydrolysates, the sardine (Sardina pilchardus) hydrolysates were characterised by the highest quantity of CGRP immunologically and biologically related molecules (Fouchereau-Peron et al. 1999). In addition, we demonstrated that using increasing hydrolysis time and various alcalase concentrations, the CGRP like molecules were mainly found after 2 hours of hydrolysis using an alcalase concentration of 0.1% (Ravallec-Plé et al. 2001). Therefore, sardine hydrolysates were prepared using these conditions and purified by gel exclusion and HPLC chromatography. The purified CGRP like molecules were then analysed for their CGRP like biological effect using the ability of CGRP to stimulate the adenylate cyclase activity in rat liver membranes. Materials and Methods Human CGRP I was obtained from Bachem. Dr D. Bataille from Montpellier, France kindly donated porcine glucagon. Labelled hormones were from Amersham (specific activity: 2000 Ci/mmole). The anti human CGRP antibody was a generous gift of Dr A. Julienne from Paris, France. Male Wistar rats were obtained from Janvier breeding, Le Genest sur Isle, France. The enzyme inhibitor Antagosan (Aprotinin- 10,000 KIU/ml) was kindly donated by Hoechst Laboratories, France. Bovine serum albumin was from Sigma and was heat-inactivated before use. All other chemicals were of reagent grade. Sardine hydrolysates were prepared from cooked head and guts using 0.1% alcalase 2.4L (Novo Nordisk Industri DK-2880 Bagsvaerd, Denmark) in phosphate buffer (0.1 M, pH 8.1, 1/10, w/v). Hydrolysis was carried out during 2 hours at 40°C. The enzyme was inactivated by 15 min of boiling in a microwave oven. After centrifugation at 25000 g, 20 min, the supernatant was filtrated through a polyethylene net, ultrafiltrated (cut off 10 KD) and freeze dried. Radioimmunoassays. Immunoreactive CGRP was measured following a previously described assay for human CGRPI (Fouchereau-Peron et al. 1990b): in brief, an anti-CGRP antiserum at a final dilution of 1/250,000 was incubated with serial dilutions of synthetic human CGRP or sardine hydrolysates. Antiserum was first incubated with tissue extracts or standard peptide for 18 h at 30°C, then 125I labelled human CGRP was added and the incubation continued during 24 hours at 4°C. Bound and free hormone was separated by charcoal- dextran precipitation. Control (specific antibody omitted) tubes were incubated in each assay. The hydrolysate was assayed in duplicate at multiple dilutions.
Liver membrane preparation. Liver membranes were obtained from male Wistar rats and prepared according to the method of Neville until step 11 (Neville, 1968). Proteins were quantified by the method of Lowry using BSA as standard (Lowry et al. 1951). Radioreceptorassays. Receptor binding ability of immunoreactive molecules was developed using rat liver membranes and 125I labelled human CGRP. Incubations, in a 400 µl final volume, were performed at 22°C during 1 hour (Yamaguchi et al. 1988a). At the end of the incubation, bound and free ligand was separated by centrifugation in a solution containing 10% sucrose. Data were expressed as specific binding that was obtained by subtracting from the total binding the amount of radioactivity associated to the membranes in the presence of 2 µM CGRP. Receptor binding ability of each purified fraction was determined in triplicate and expressed as the quantity of protein (µg) that induced a 25% inhibition of the initial binding. Adenylate cyclase activity. The adenylate cyclase activity in rat liver membranes was determined by measuring the synthesis of cAMP from non radioactive ATP as previously described (Yamaguchi et al. 1988b). The assay was initiated by addition of membranes (12 µg) to preheat (4 min at 30°C) assay tubes. Membranes were incubated for 30 min at 30°C in 50 µl Hepes Tris buffer (25 mM, pH 7.4) containing 1 mM isobutylmethylxanthine (IBMX), 1.3 mM EGTA, 5 mM MgCl2, 0.4 mg/ml BSA, 1 mM ATP and 100 µM GTP. The buffer also contained 10 mM phosphocreatine and 0.8 mg/ml creatine phosphokinase as regenerating system. After incubation, the reaction was terminated by the addition of 200 µl Tris –EDTA buffer, pH 7.4 and the tubes placed in a boiling water bath for 4 minutes. Cyclic AMP was quantified on 50 µL aliquots using the radioreceptor assay kit from Amersham. The adenylate cyclase activity was expressed as picomoles of cAMP synthesised by 1 mg membrane protein during 1 min of incubation. The adenylate cyclase activity was also measured in conditions specifically used to demonstrate a glucagon stimulated adenylate cyclase activity (Hanoune et al. 1977). 20 µg of liver membranes were incubated for 10 min at 30°C in 50 µl Tris HCl buffer (50 mM, pH 7.6) containing 1 mM isobutylmethylxanthine (IBMX), 1 mM EDTA, 3 mM MgCl2, 0.9 mg/ml BSA, 0.09 mg/ml bacitracin, 0.5 mM ATP and 1 µM GTP. The buffer also contained 25 mM phosphocreatine and 1 mg/ml creatine phosphokinase as regenerating system. After incubation, the experiment was processed as previously described for the CGRP stimulated adenylate cyclase activity.
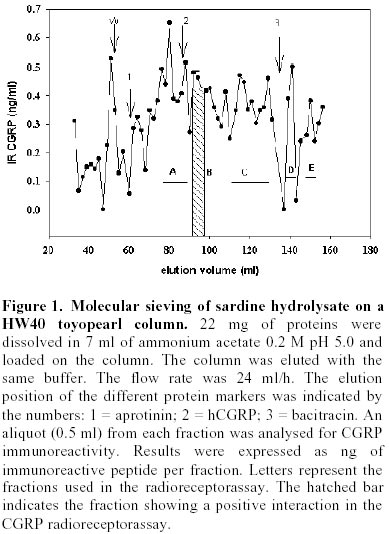
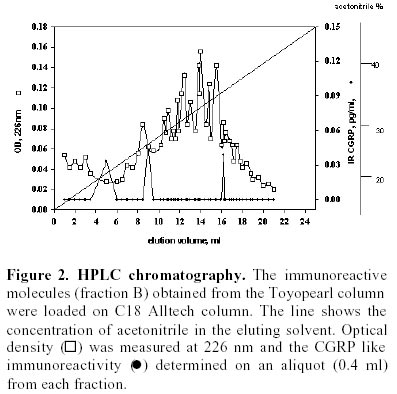
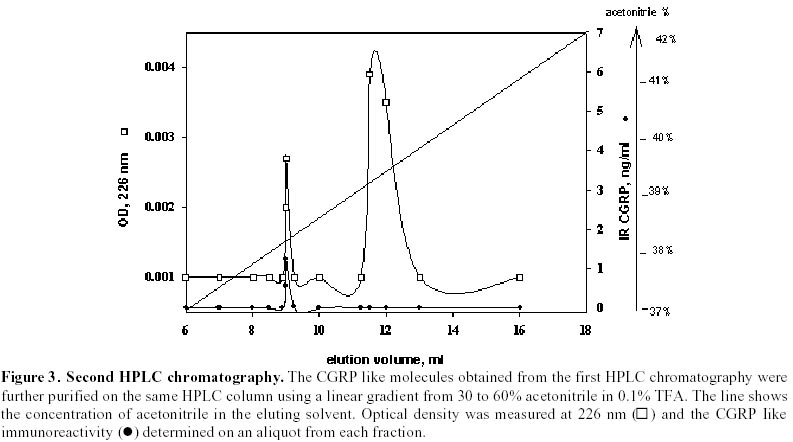
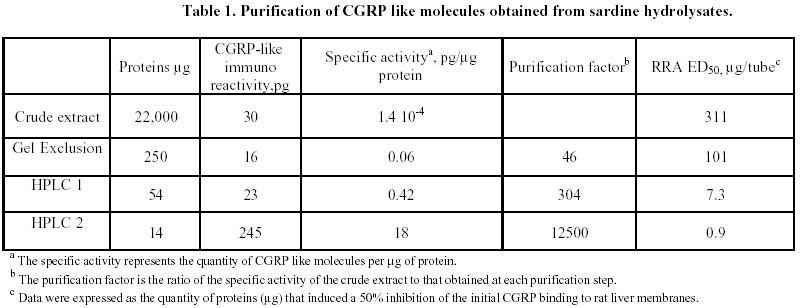
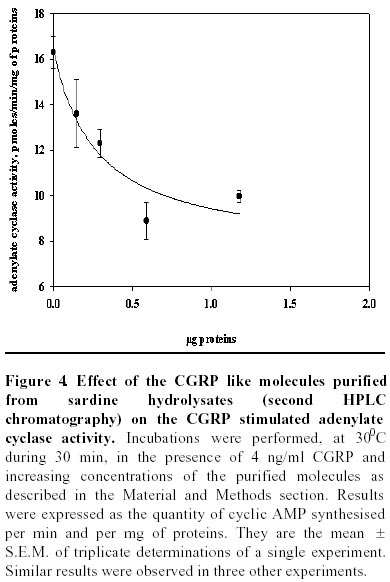
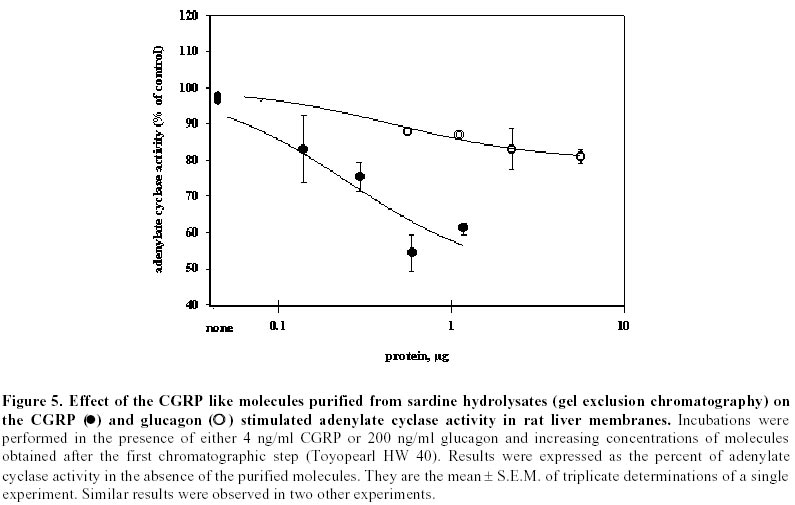
Purification of sardine hydrolysates. Sardine hydrolysates were prepurified by gel exclusion chromatography on a HW 40 toyopearl column (2.5 x 33.5 cm) using ammonium acetate 0.2 M, pH 5 as eluant. The flow rate was 24 ml/hour. Immunoreactive fractions were analysed using CGRP radioreceptorassay. Positive fractions were then subjected to HPLC on a C18 prosphere column using a linear gradient of 10 to 60% acetonitrile in 0.1% TFA. The biological activity of these fractions was measured by radioreceptorassay and positive fractions were then repurified using the same column and a linear gradient of 30 to 60% acetonitrile in 0.1% TFA. Optical density was measured at 226 nm. Mass spectrophotometry was performed by L. Marvin from University of Rouen. Protein determination. The protein concentration of each analysed fraction was performed using the bicinchoninic acid method using BSA as the standard (Smith et al. 1985). Results Purification of sardine hydrolysates Sardine hydrolysate (22 mg of protein suspended in 7 ml of eluant) was loaded on the HW40 toyopearl column. 2 ml fractions were collected and a 0.5 ml aliquot was used to quantify the CGRP immunoreactivity. The CGRP immunoreactive profiles of this preparation (Figure 1) demonstrated 5 main immunoreactive fractions (referred as A, B, C, D and E). These fractions were further analysed using the CGRP radioreceptorassay. Only the second tested fraction was able to inhibit the labelled CGRP binding as the unlabeled hormone did (data not shown). Fraction B was further purified by HPLC chromatography (Figure 2). 250 µg of proteins were loaded on the column, 1 ml fractions were collected and a 0.4 ml aliquot used to determine the CGRP immunoreactivity. Immunoreactive profile of this separation gave three main immunoreactive peaks. These immunoreactive fractions were tested for their ability to inhibit the 125I CGRP binding to rat liver membranes. The data showed that the most efficient inhibiting effect resulted with the immunoreactive material present in the third peak. Fifty percent inhibition was observed with 13 µg of proteins from this peak. This fraction eluted for an acetonitrile concentration of 35%. In the same conditions, human CGRP eluted for 33.5% acetonitrile. 53 µg proteins of this fraction were subjected to a second HPLC chromatography using the same column and a linear acetonitrile gradient from 30 to 60%. Two absorbency peaks were observed (Figure 3): Only the fraction 9 showed CGRP -like immunoreactivity. In the CGRP radioreceptorassay, this fraction displaced the labelled CGRP binding: 25% inhibition was observed with 370 ng of proteins, that is a K0.5 of 6.5 x 106M-1. No significant displacement was observed with the second peak. So, from 22 mg of sardine hydrolysate proteins (Table 1) we obtained 14 µg of a molecule that present CGRP biological and immunological activity. The purification factor obtained was about 12.500. The different purification steps were followed using the CGRP radioreceptorassay. While 311 µg of proteins from crude extract were necessary to displace 50% of the initial CGRP binding to rat liver membranes, only 0.9 µg of proteins from the final purification step produced the same inhibitory effect (Table 1). The molecular weight of this molecule determined by mass spectrophotometry was 6000 daltons. Adenylate cyclase activity of the purified fraction The biological activity of the fraction obtained from the second HPLC was analysed by referring to the capacity of CGRP to stimulate the adenylate cyclase activity in rat liver membranes. The effect of this fraction was analysed on the CGRP stimulated adenylate cyclase activity (Figure 4). Increasing protein concentrations between 0.15 and 1.2 µg of proteins induced a dose related inhibition of the stimulated activity. 25% inhibition was observed with 0.3 µg of proteins, that is a K0.5 of 106M-1. In order to analyse the specificity of the observed inhibition, we compared the effect of the fraction purified after gel exclusion chromatography on the CGRP (4 ng/ml)- and glucagon (200 ng/ml) stimulated adenylate cyclase activity in the same rat liver membrane preparation (Figure 5). The used glucagon concentration represents the dose inducing the maximal response in rat liver membranes (Hanoune et al. 1977). At these concentrations, the stimulated adenylate cyclase activity represented a two and twenty fold increase over the control (8.4 ± 0.4 and 11 ± 1 pmoles of cyclic AMP per min and per mg of proteins) for CGRP and glucagon, respectively. The inhibiting effect of the prepurified fraction was significant only on the CGRP stimulated adenylate cyclase activity. In the presence of 1.1 µg of proteins, the inhibiting effect observed was 14 and 35% of the glucagon and CGRP stimulated adenylate cyclase activity, respectively. The same protein concentration induced a 10% inhibition of the liver membrane adenylate cyclase activity. Discussion Many bioactive peptides have been isolated from enzymatic hydrolysates of proteins (Hsueh and Moskowitz, 1973; Piot et al. 1992; Nakagomi et al. 2000). Indeed, invertebrate and fish by-products might be the best source for such active biopeptides because these substances would make good starting materials for safer and less toxic medicines. In addition, these bioactive peptides may be used as adjuvants to stimulate food intake and to enhance growth and disease resistance of animals We reported here the purification from sardine hydrolysates of CGRP related molecules. Sardine hydrolysates prepared using specific conditions, that is two hours of hydrolysis with 0.1% alcalase were subjected to purification by size exclusion chromatography and HPLC. The appearance of the CGRP related molecules was followed using both radioimmunoassays and radioreceptorassays. From 22 mg of proteins present in the crude extract we obtained 14 µg of CGRP related molecules. These molecules were obtained with a purification factor of 12500 and present a molecular weight of 6000 daltons. This molecular weight is identical to the apparent molecular weight we reported after exclusion chromatography of sardine hydrolysates prepared using a different condition that is four hours of hydrolysis with 1% alcalase (Fouchereau-Peron et al. 1999). In order to analyse the function of these molecules, we tested their ability to modulate the adenylate cyclase activity in rat liver membranes: a specific target tissue for CGRP. These experiments show that they have a specific inhibitory action on the CGRP stimulated adenylate cyclase activity. No such inhibitory effect was observed on the glucagon stimulated adenylate cyclase activity of the same membrane preparation. The same results were observed using hydrolysates prepared using different conditions that is after 4 hours of hydrolysis with 0.1% alcalase (data not shown). So, whatever the conditions of hydrolysis, the use of alcalase allows to obtain CGRP related molecules whose molecular weight is around 6000 daltons and that present an inhibitory action on the CGRP induced adenylate cyclase activity in rat liver membranes. So these purified molecules are able to decrease the 125I labelled CGRP binding to rat liver membranes and to inhibit the following biological effect. This action both at the receptor and the adenylate cyclase level is similar to that observed using a well known competitive inhibitor of CGRP action: the human CGRP 8-37 (Chiba et al. 1989). 0.3 µg of our purified proteins, that is 1 10-6M, induced a 25% inhibition of the CGRP stimulated adenylate cyclase activity. In the same conditions and using the same rat liver membrane preparation, the effect of 10-6M CGRP 8-37 induced the same inhibiting effect. Thus, our purified CGRP related proteins appear at least as efficient as CGRP8-37 a specific antagonist of CGRP action in rat liver membranes. So, these purified molecules may act as antagonists for peptides that bind to CGRP receptors in rat liver membranes. In addition, they are probably C terminal fragments as all CGRP antagonists which were reported belong to this class of molecules (Rist et al. 1998). So, the enzymatic hydrolysis of sardine by-products allows obtaining molecules immunologically and biologically related to CGRP. The presence of these molecules in fish hydrolysates is not surprising as immunoreactive CGRP like molecules are mainly found in gill and intestine of non mammalian vertebrates (Fouchereau-Peron et al. 1990b). In non-mammalian vertebrates, this peptide, by its action on the gill, controls hydromineral metabolism. In addition, the high circulating levels of CGRP in teleosts suggests that this neuropeptide is indispensable for everyday functions such as food intake in teleosts. Recent data demostrated a direct relationship between the CT and calcium plasma levels in eels after a high calcium diet (Suzuki et al. 1999). The development of new antagonists may be of particular importance in various aspects of the CGRP action mainly in the control of feeding where CGRP together with amylin exert a direct or indirect control of meal size and the control of meal initiation (Geary, 1999). References
Supported by UNESCO / MIRCEN network © 2001 by Universidad Católica de Valparaíso -- Chile The following images related to this document are available:Photo images[ej01004f2.jpg] [ej01004t1.jpg] [ej01004f3.jpg] [ej01004f1.jpg] [ej01004f4.jpg] [ej01004f5.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}