 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Electronic Journal of Biotechnology, Vol. 4, No. 1, April, 2001 SHORT COMMUNICATION Nucleotide sequence of a genomic clone encoding a cowpea (Vigna unguiculata L.) trypsin inhibitor Paulraj Kirubakaran Lawrence*1, Jayaveeramuthu Nirmala2 and Kripa Ram Koundal3 1Department
of Biological Sciences, Texas Tech University, Box No 43131, Lubbock, TX 79409-3131
USA Tel: 806-742-2740 Fax: 806-742-2963 E-mail: p.k.lawrence@ttu.edu
Financial support: Senior Research Fellowship provided to PKL, by Council for Scientific and Industrial Research, New Delhi, India-110012. Received
December 15, 2000 Code Number: ej01006 Abstract A protease inhibitor gene has been isolated from native cowpea (Vigna unguiculata L.) cv. 130 from l-ZAP II genomic library. The nucleotide sequence of this genomic clone shared 86% homology with a cowpea trypsin inhibitor f IV mRNA, and 81% with Bowman-Birk protease inhibitor genes of soybean. The isolated gene had TATA and CAT signals in the 5' upstream region. The longest open reading frame had 504 bases, encoding 167 amino acids from the predicted coding region. The active serine centers of this protein was found between 105-158 amino acid residues, with a 69 amino acids long signal peptide in its N terminal. Keywords: cowpea genomic library, nucleotide sequencing, sequence analysis. Article Proteinase inhibitors are potential tools of crop improvement targeting plant protection and human nutrition. This concept of plant protection is rapidly expanding and opens up diverse areas to harness the potential of these genes. The serine and its trypsin subclass of protease inhibitors exhibit antimetabolic and antinutritional effects against a wide variety of crop pests belonging to the orders of Lepidoptera, Coleoptera and Orthoptera (Shulke and Murdock, 1983). These inhibitors are lethal to the larvae of Callosobruchus maculatus (Gatehouse and Boulter, 1983) and Manduca sexta (Shulke and Murdock, 1983) at 10% of their diet. They also promote the loss of sulphur based amino acids, leading to weak and stunted growth and ultimate death. These inhibitors are cysteine rich with a molecular mass of 8-20 kD (Peng-Wen Chen et al. 1997). Many of these are products of multigene families and exhibit different specifications towards proteases (Ryan, 1990).
The gene size and coding regions of these inhibitors are small, devoid of introns (Boulter, 1993), and comprised of readily identifiable core region covering the invariant cysteine residues and active serine centers that are bound to highly variable amino and carboxy terminal regions. The reaction sites of these inhibitors are mutating faster than amino acids in rest of the proteins, implying their role in defense against insect pests, which may exert a strong selection pressure on these proteins to conserve the reaction sites related to plant defense (Laskowski et al. 1988). The availability of these inhibitors may allow to engineer transgenic crops inherently resistant to these insect pests. These aspects therefore encourages the identification and characterization of novel protease inhibitor genes from diverse plant species to be used against insect pest. This report describes the screening of a cowpea genomic library with a tomato protease inhibitor cDNA probe to identify, characterize and clone a new protease inhibitor gene. Materials and Methods Construction and screening of genomic library. Genomic DNA of one week old etiolated seedlings of cowpea (Vigna unguiculata L.) cv. V130 was isolated (Dellaporta et al. 1983), purified and partially restricted with EcoRI. This was fractionated on a sucrose density (10-30%) gradient and fragments of 2-7 kb were pooled together and approximately 10 mg of DNA was used for cloning in l-ZAP II vector following the manufacturer's protocol (Stratagene, La Jolla, USA) and amplified in E. coli XL-1 Blue. The library was screened using a 700 bp cDNA clone encoding tomato protease inhibitor II (pT2, obtained from Prof. Ryan, Washington State University, Pullman, USA) according to the method of Sambrook et al. (1989). Positive clones were identified after 3 rounds of screening.
In vivo excision and sequencing. The 3 recombinant lambda clones obtained after tertiary screening were excised into pBluescript II SK+ plasmid using ExAssist helper phage (Short et al. 1988) and transformed into SOLR (Stratagene, La Jolla, USA) super competent cells. DNA from 10 excised positive clones (pLK1 to pLK10) was isolated using Qiagen mini-prep kit (Qiagen Inc., Valencia, USA) and digested with EcoRI (5 U/mg of DNA) in a volume of 20 ml. The digested plasmid DNA was analyzed on 0.7% agarose gel, transferred to Hybond N+ membrane and probed using pT2 (Sambrook et al. 1989).
One of the genomic clone (pLK8) was subjected to exonuclease III deletion using MBI Exo III kit (MBI Fermentas, Hanover, USA) and transformed into E.coli DH5a . The plasmid DNA (3-4 mg) from ten successive overlapping clones were sequenced by di-deoxy chain termination method (Sanger et al. 1977) using Sequenase ver 2.0 sequencing kit (Amersham Inc., Cleveland, USA). The clones were sequenced using both forward and reverse M13 universal primers. The reaction products were separated on 8% polyacrylamide gel containing 8 M urea using LKB Macrophor (LKB Pharmacia, Switzerland) equipment. The nucleotide sequence was analyzed by Lasergene programme (DNASTAR), Genbank BLASTN (Altschul et al. 1997) and FASTA (Pearson and Lipman, 1988).
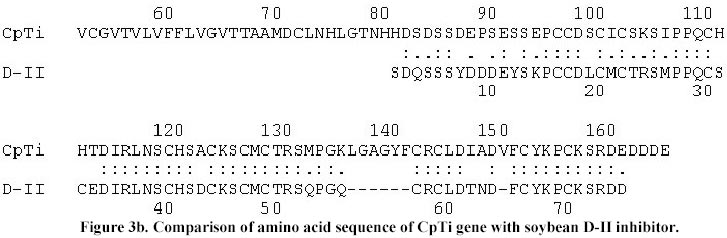
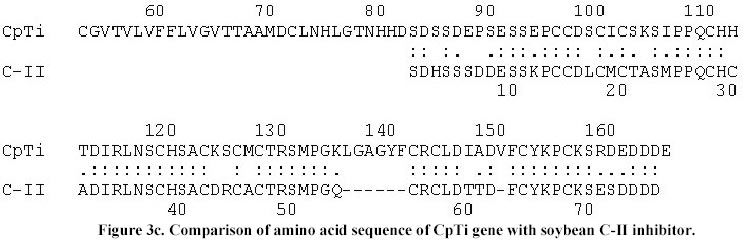
The sequence data has been submitted to EMBL nucleotide data base (Accession No. AJ 271752). Results and Discussion After 3 rounds of screening 3 positive lambda clones were obtained. In vivo excision resulted in 10 plasmid clones, all having an insert size of 2 kb when restricted with EcoRI. Hence, one of the genomic clone designated as pLK8 was sequenced after deletion with Exonuclease III. The deletion of genomic clone pLK8 resulted in successive overlapping clones of approximately 200-225 bp shorter, ranging from 2-1.8 kb. These clones had approximately 25 bp region overlapping with the preceding clone. These clones were sequenced using forward and reverse primers in order to confirm sequences from both DNA strands. The sequences of deleted clones were aligned (Megalign) and its total length was found to be 1997 bp. The sequence from 1-990 bp of the original 2 kb clone did not show any significant homology with any protease inhibitor gene or its promoters, so 1007 bp are shown in Figure 1. The clone had transcription control regions like TATA signals between 256-267 bases and a CAT signal at 335 bp. The longest ORF analyzed was between 355 and 858 bases in the first reading frame and the ATG for this ORF starting at 355 bp. This ORF on translation gave a protein with 167 amino acids (indicated by one letter code, Figure 1). The molecular weight of this protein was calculated to be 18.5 kD; of which 18 amino acids were strongly basic, 20 strongly acidic, 40 hydrophobic and 62 are polar amino acids with an isoelectric point of 6.5. The protein has a N terminal signal sequence of 69 amino acids which is important in its translocation as has been reported in many proteinase inhibitors such as soybean (Hammond et al. 1984), cowpea (Hilder et al. 1989), pea (Domoney et al. 1995), maize (Rohrmeier and Lehle, 1993) and alfalfa (McGurl et al. 1995). The cowpea protease inhibitor has the signal peptide clevage site between 69-70 amino acid residues i.e., TTA-AM, which is comparable to the alfalfa trypsin inhibitor (ATI) having a signal peptide of 44 or 55 amino acid residues which may target the ATI to the ER and then eventually deposit it in the vacuole of the cell (McGurl et al. 1995). Comparison for conserved domain in the protein by NCBI CD-search showed that the regions from 105-158 have the active domain similar to serine class of Bowman-Birk inhibitors. The active centre serines (6) are located with in this above region. The total number of cysteine residues (15) are identical to that of the cowpea f IV inhibitor mRNA (Hilder et al. 1989) with most of them found in the active domain. The total number of bases translated in the ORF is 504, with 58.28% A+T and 41.72% C+G similar to that of cowpea trypsin inhibitor mRNA f IV (Hilder et al. 1989) which has 60.5% A+T and 39.4% G+C in their expressed regions. The regions from 627-851 bp of the sequenced fragment shared 81% homology with 191-394 bp of soybean C-II protease inhibitor gene which is 645 bp long (Jourdrier et al. 1987), and with 138-341 bp of BBI mRNA (Baek et al. 1994), which is 488 bp long respectively. The maximum percentage (86%) of homology (Figure 2) was found with cowpea trypsin inhibitor mRNA coding for inhibitor f IV which is 583 bp long (Hilder et al. 1989). Further, it was observed that sequences between 627 to 857 shared homology with the coding regions of all the above mentioned inhibitors. The protease inhibitor genes are typically devoid of any intervening sequences (Boulter, 1993) and none of the genes sequenced so far have any introns, similarly the isolated sequenced cowpea trypsin inhibitor gene is also devoid of introns. The amino acid sequence of this trypsin inhibitor shared a 80% homology (Figure 3a) with the protein of cowpea trypsin inhibitor f IV, 63.4% with soybean D-II inhibitor (Figure 3b) and 65% with soybean C-II inhibitor (Figure 3c). From the above analysis it could be clearly concluded that the isolated genomic clone had the complete gene encoding a trypsin inhibitor and designated as CpTi, which could be used as a potential tool against crop pests. This gene has been cloned in Bin AR, a binary vector and used to transform pigeonpea by Agrobacterium tumefaciens mediated genetic transformation. The integration of this gene in transformed pigeonpea has been confirmed by Southern hybridization and also its expression by northern analysis (Lawrence and Koundal, 2001). These transformed plants are further to be tested against pod borer (Heliothis armigera) which is under progress. Acknowledgements We are thankful to Prof. C.A. Ryan Washington State University, Pullman, for providing tomato protease inhibitor cDNA probe. One of the authors (P.K.L.) also wish to thank the Council of Scientific and Industrial Research for granting fellowship during the course of this investigation. References
Supported by UNESCO / MIRCEN network © 2001 by Universidad Católica de Valparaíso -- Chile The following images related to this document are available:Photo images[ej01006f3b.jpg] [ej01006f3a.jpg] [ej01006f2.jpg] [ej01006f1.jpg] [ej01006f3c.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}