 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Electronic Journal of Biotechnology, Vol. 4, No. 3, December, 2001 Analysis of Uruguayan weedy rice genetic diversity using AFLP molecular markers María Teresa Federici*1, Duncan Vaughan2, Norihiko Tomooka3, Akita Kaga4, Xin Wang Wang5, Koji Doi6, Marta Francis7, Gonzalo Zorrilla8 and Nestor Saldain9 1Unit
of Biotechnology, National Institute of Agronomical Research, Experimental Station
"Las Brujas", Ruta 48. Km. 10. Rincón del Colorado. Canelones,
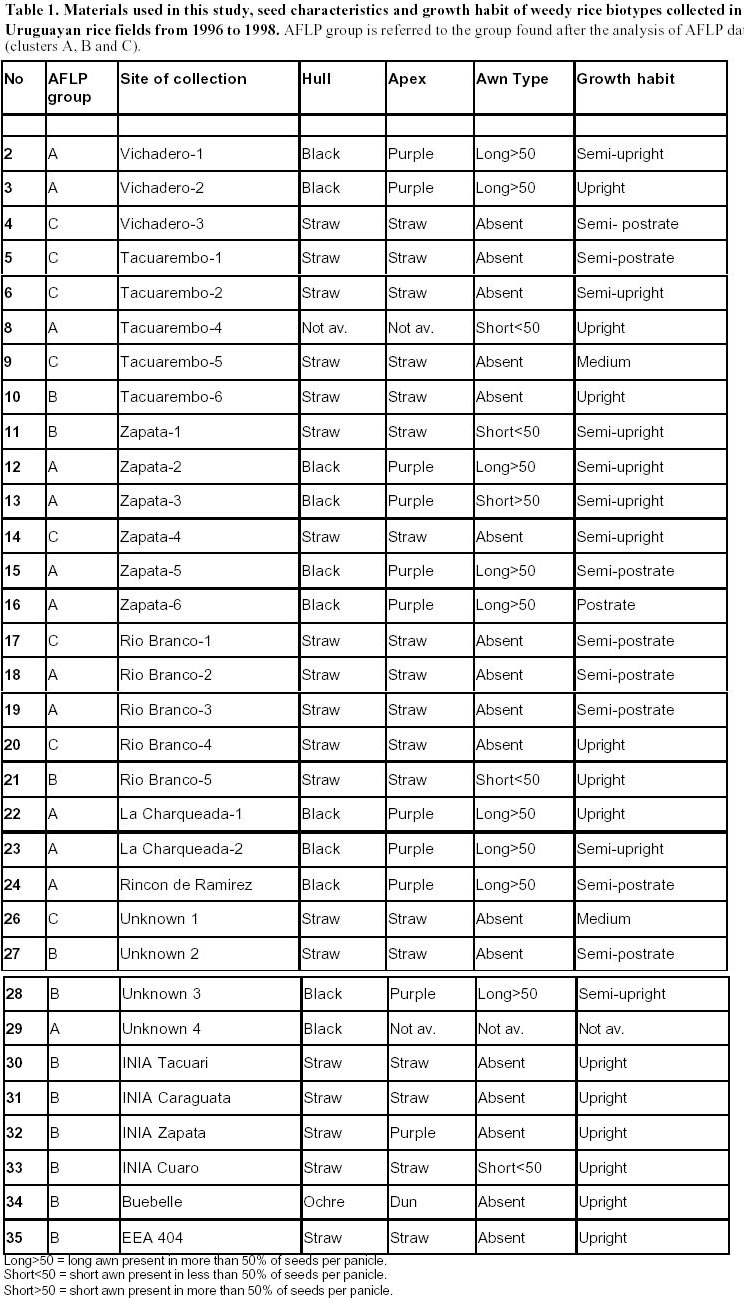
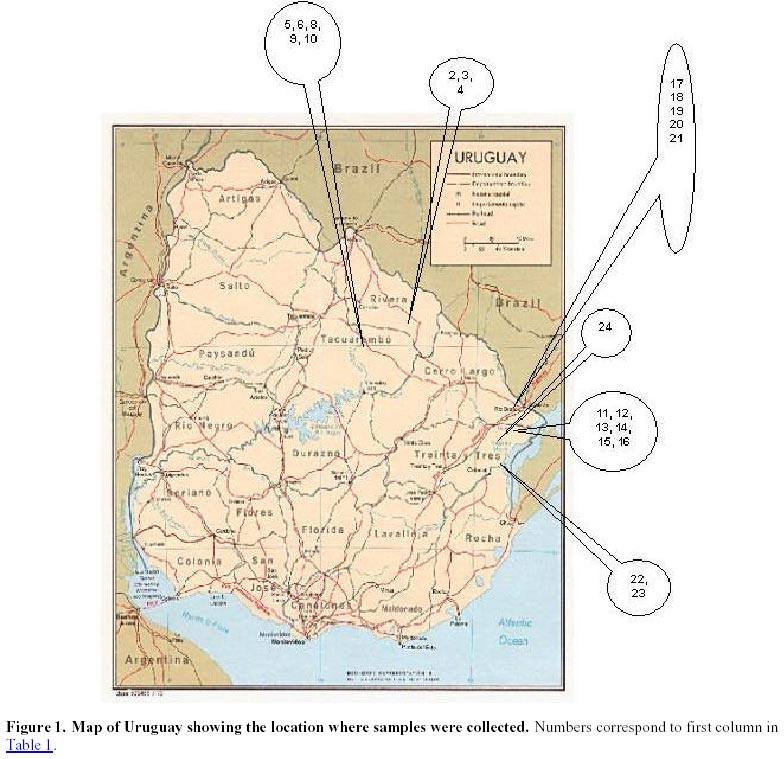
Uruguay Tel: 598-2-0327-7641 Fax: 598-2-0327-7609 E- mail: maritefe@inia.org.uy Financial support: Japan Cooperation Agency (JICA)- Individual Training. Received June 22, 2001 Code Number: ej01026 Abstract Weedy rice is a serious problem in Uruguayan rice fields since intensification of rice production started about 10 years ago. The genetic diversity of 26 weedy accessions of weedy rice and 6 Uruguayan cultivars were analyzed using AFLP (amplified fragment length polymorphisms) methodology. Abundant polymorphisms were found among samples tested. Using different methods of analysis three groups of samples were revealed. A relationship was found between three groups and morphological traits. One group had a black hull, purple apex and long awn (wild type traits) while another group had straw hull and apex, and short or no awn (domestication traits). The third group included the cultivars analyzed and some weedy rice samples. The weedy rice in this third group is presumed to most closely mimic cultivated rice and may have recently evolved. The results suggest that weedy rice adapts either to the natural environment or to cultivation. The former type with black hull and long awn may be easy to control because it can easily be seen. The later group may be difficult to control, particularly since the weedy rices within the cluster consisting of cultivars suggest that weedy rices are continually evolving in Uruguayan rice fields. The AFLP technique is very effective for assessing genetic diversity within weedy rice and will be very useful for fingerprinting of local cultivars of rice. Keywords: clusters, dendrograms, polymorphic bands, red rice, population dynamics. Article Red rice has always occurred in Uruguayan rice fields at low population levels. Over the last 10 years, the intercropping period was reduced due to more intensive rice production. At the same time, red rice populations and the number of infested rice fields have increased becoming a serious rice production problem. The term "weedy rice" refers to populations of usually annual Oryza species that diminish farmer income both quantitatively through yield reduction, and qualitatively, through lowered commodity value at harvest. Weedy rice occurs in all the major rice growing areas in the tropics, being a particular problem in direct seeded rice agriculture of Latin and North America, the Caribbean, Africa, South and South East Asia (Oka, 1988). Weedy rice occurs in temperate regions where no wild relatives of rice occur in these areas, such as the USA and Europe. Weedy rice occurs where conditions of direct seeding and intensive production prevail (Bres-Patry et al. 2001). Weedy rice may have indica or japonica characteristics and show weedy traits such as phenotypic plasticity, a high seed dispersal ability and seed dormancy. Weedy rice seeds usually have a red pericarp (thus commonly called "red rice"), earlier tillering plus flowering than cultivated rice, and anthocyanin pigmentation of different plant parts, such as, collar, ligule, grain apiculus, stigma and awns (Cho et al. 1995; Suh et al. 1997). Several studies on weedy rice genetic diversity have been reported. Weedy rice strains also appear to be differentiated into indica and japonica types based on morphological and physiological traits, isozymes, restriction fragment length polymorphism (RFLP) and random amplified polymorphic DNA (RAPD) markers (Cho et al. 1995; Suh et al. 1997). DNA fingerprinting using simple sequence repeats (SSR or microsatellites) has also been reported indicating differentiation among weedy rice populations (awnless types; straw hull and black hull awned types; cultivated rice and hybrids between cultivars and each weedy type respectively (Gealy, 2000, personal communication). Recently, amplified fragment length polymorphism (AFLP) methodology has been found to be a useful tool to study relationships within members of various taxa (i.e. Caicedo et al. 1999; Yoon et al. 2000). The AFLP technique is a novel polymerase chain reaction (PCR)- based molecular marker assay (Vos et al. 1995) that has the capacity to detect a higher number of loci and thus a higher rate of polymorphism in a single assay than RFLPs or RAPDs (Powell et al. 1996). AFLP methodology has been employed, because of their effectiveness and reliability for studies on diversity, phylogeny, genomic linkage mapping, and identification of varieties (Becker et al. 1995; Maughan et al. 1996; Xu et al. 2000). In this study genetic diversity among weedy and cultivated rice was assessed using the AFLP technique. The objectives of the study were to use the AFLP technique to fingerprint 26 biotypes of weedy rice and 6 cultivars from Uruguay and analyze genetic diversity in these materials; determine genetic relationships between weedy rice and Uruguayan cultivars and find out if this provides insights into the origin of weedy rice; and relate diversity analysis to strategies to control weedy rice in Uruguayan fields. Materials and Methods Plant materials. Panicles of different red rice biotypes were collected from rice fields across Uruguay from 1996 to 1998 (Table 1, Figure 1). Individual plants of each type were transplanted in the field in 1999. Records of morphological traits used as variety descriptors were taken on a single plant basis. In March 2000, seeds were harvested from these plants. The seeds were grown in the tropical greenhouse of NIAR, Tsukuba, Japan for DNA extraction. DNA analysis. Total genomic DNA was extracted from young leaves of 20 days old plants using the CTAB method (Saghai-Maroof et al. 1984). Briefly, 3 g of fresh leaf tissue were finely ground after immersion in liquid nitrogen (at this step material can be stored –80ºC). 10% CTAB solution containing 0.2% beta-mercaptoethanol, was added to the finely ground leaves and incubated at 65ºC in a water bath with shaking for 20 min. The solution was mixed with 20 ml. of chloroform/ isoamylalcohol (24:1), mixed by inversion at room temperature during 20 min, and then centrifuged at 5000 rpm at RT during another 20 min. The supernatant was then mixed with 2 ml of pre- heated 10% CTAB solution and incubated for 3-5 min at 65ºC. After that, an equal volume of chloroform /isoamylalcohol was added, mixed at room temperature for 20 min and centrifuged at 5000 rpm during 20 min (at room temperature). Then, 1.5 volumes of CTAB precipitation buffer (1% CTAB, 5 mM Tris HCl, 10 mM EDTA pH 8.0) was added and fully mixed. After storing at room temperature 10 min, the mixture was centrifuged at 3000 rpm for 20 min (also at room temperature). After that, samples were incubated with 10 ml of buffer (1M NaCl, 10 mM Tris HCl, 1 mM EDTA) at 65ºC to dissolve pellet completely and treated with 8 to 10 ml of RNAse (810 mg/ml) at 37ºC during 30 min. Then, the enzyme was inactivated by incubating samples at 65ºC during 10 min. After ethanol precipitation, the pellet was washed twice with 70% ethanol during 7 min followed by another 5 min wash with 99.5% ethanol, dissolved in 0.1X TE and stored at –20ºC for use. DNA concentrations were estimated in 1% agarose gels and adjusted to 50 ng/µl. PCRs were performed as described by Vos et al. (1995) with minor modifications. The procedures followed were: 500 ng of DNA from each sample were digested with 5 units of EcoRI (recognition sequence 5`GAATTC-3`) and 5 units of Mse I (5’ TTAA 3’) in a final volume of 40 µl. This mixture was incubated at 37ºC during 3:30 hours. The reaction buffer was 5X RL-buffer (50 mM Tris. HAc pH 7.5, 50 mM MgAc, 250 mM KAc, 25 mM DTT, 250 ng/µl BSA). To the double digested DNA sample, a total volume of 10 µL of ligation reaction mixture including 5 pmoles of EcoRI adapter and 50 pmoles of Mse I adaptor, 10 mM ATP, 1 unit of T4-DNA ligase, and the same reaction buffer as the above mentioned were added and then incubated overnight at 37ºC. Adapter sequences used are listed (Table 2). Digested/ ligated DNA samples were diluted 10 times with TE. A 20 µl pre-selective PCR mixture contained 5 µl of diluted template mixture, 75 ng of both MseI 00 primer (MseI adaptor sequence) and EcoRI 00 primer (EcoRI adaptor (Table 2), 200 µM dNTPs, 10 mM Tris-HCl (pH 8.3), 1.5 mM MgCl2, 50 mM KCl, and 1 unit Taq polymerase. The cycle profile used was: 94ºC 30 sec, 65ºC 30 sec, 72ºC 60 sec (cycle 1), then annealing temperature is lowered 0.7ºC each cycle during next 12 cycles and 94ºC 30 sec, 56ºC 30 sec and 72ºC 60 sec from cycles 14 to 36. The pre-amplification product was diluted 20 times with TE (10 mM Tris HCl, pH 8.0, 0.1 mM and EDTA- Etylene-diamine-tetra-acetic acid), and then used as a template for selective amplification to generate AFLPs. The selective amplification reaction was conducted in a final volume of 20 µl containing 5 µL of diluted preamplification reaction, 30 ng of each MseI+3 and EcoRI+3 primers (Table 2), 200 µM dNTPs, 10 mM Tris-HCl (pH 8.3), 1,5 mM MgCl2, 50 mM KCl, and 1 unit Taq polymerase. Amplifications were conducted on a 9600 Perkin Elmer DNA thermocycler (Norwalk, CT). The PCR was programmed with the same cycle profile above mentioned.A total of 5 µl of the AFLP selective amplification product were mixed with 2.5 µl of loading buffer (98% formamide, 10 mM EDTA, 0,25% each of bromophenol blue and xylene cyanol FF), then denatured at 95ºC for 2 min and place immediately on ice. Electrophoresis was performed on an EC 160 standard sequencing unit (E-C Apparatus Corporation, USA). About 3 µl of mixture were loaded onto 6% (w/v) polyacrylamide denaturing gel with 1X TBE buffer after a pre-run electrophoresis at 50 W for 1 hour, and then were run at 50 W for about 2 hours. After electrophoresis, the gel was silver- stained using the silver sequence DNA staining kit (Promega cat # Q4132, Madison, WI) following the instructions provided. AFLP bands were visually scored with the aid of digital pictures of the gels and adobe Photoshop computer software. AFLP bands were scored as present (1) and absent (0) and only bands showing unambiguous polymorphism were entered into a data matrix that was used for the following analyses. Genetic similarity was calculated with the simple matching coefficients (Sokal and Michener, 1958) by the NTSYS-pc version 1.70 computer software (Rohlf, 2000). Then, the similarity matrices were subjected to cluster analysis by the unweighted pair-group method with the arithmetic averages (UPGMA) cluster method and Plus analysis (Innan et al. 1999). Results
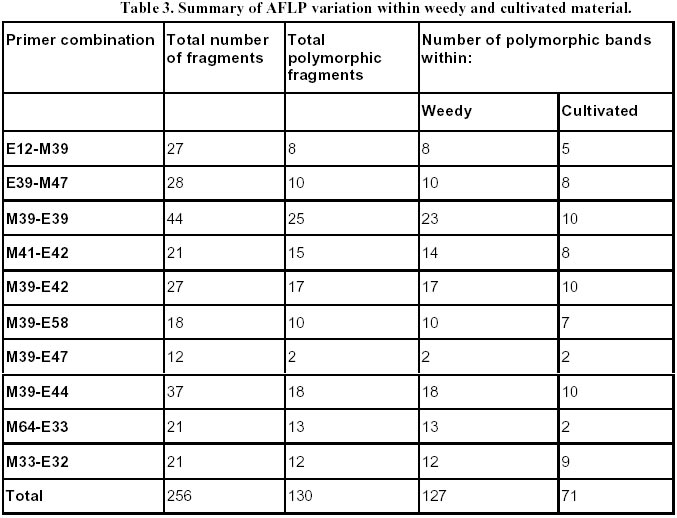
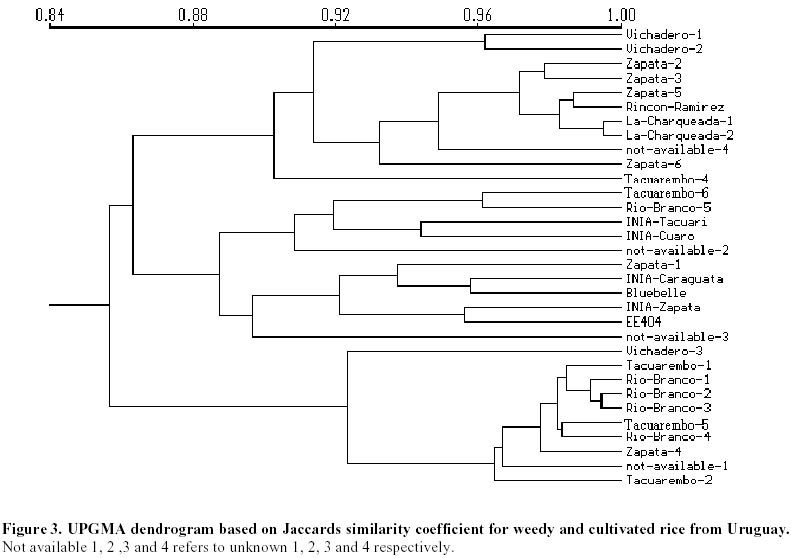
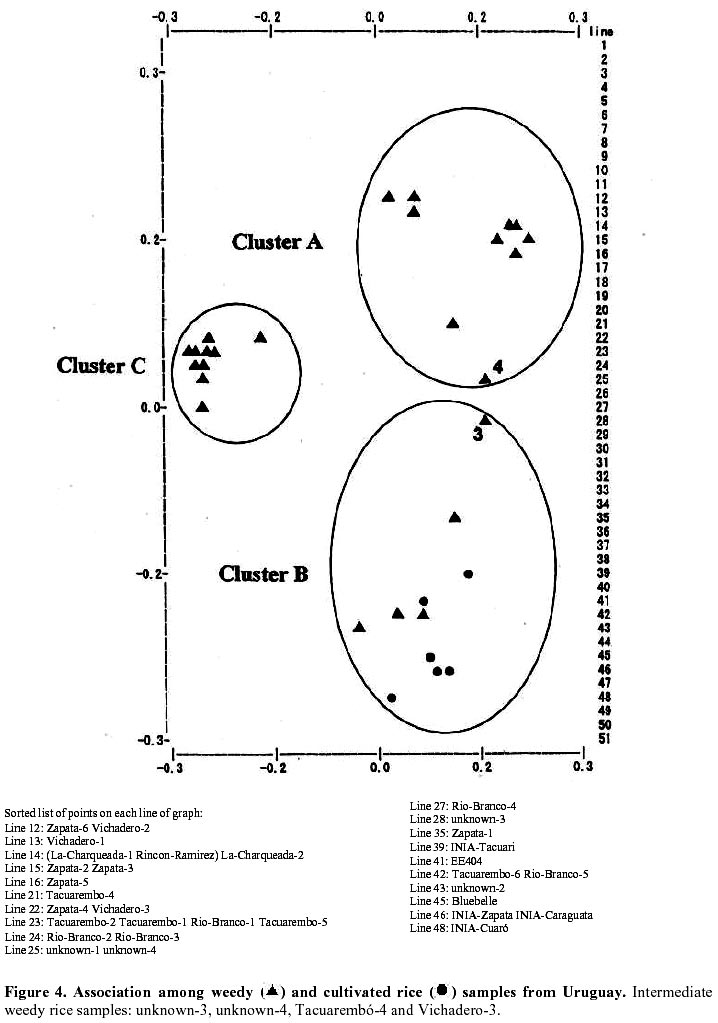
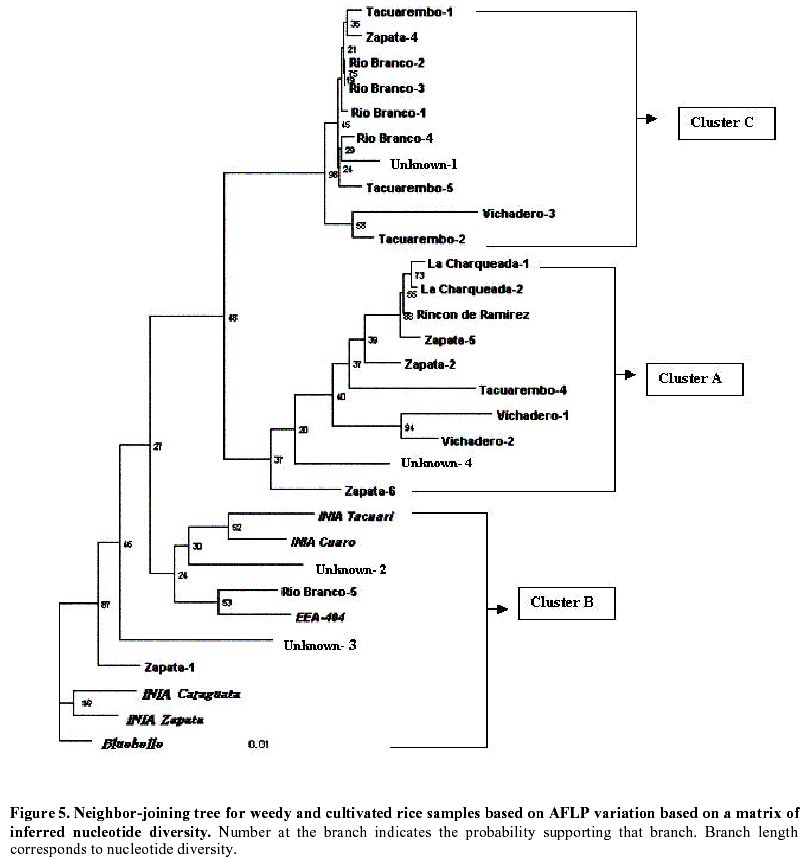
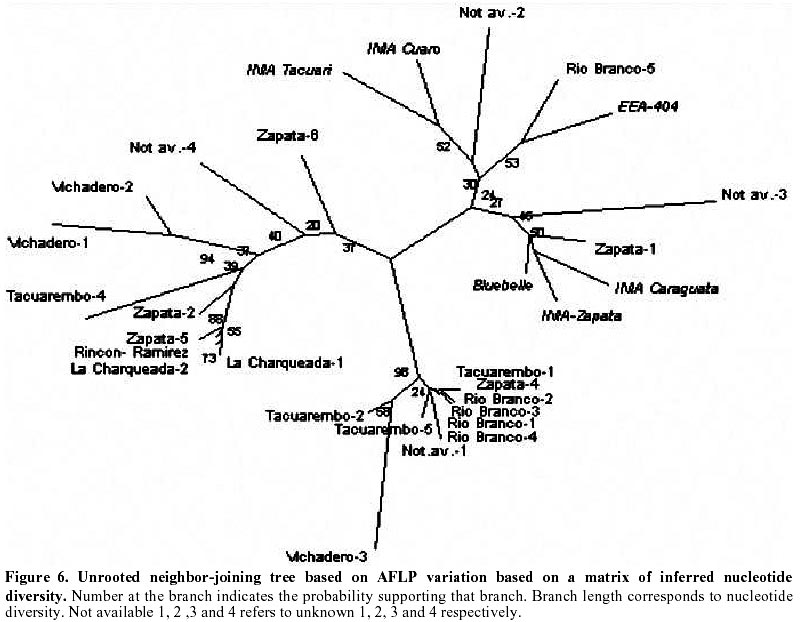
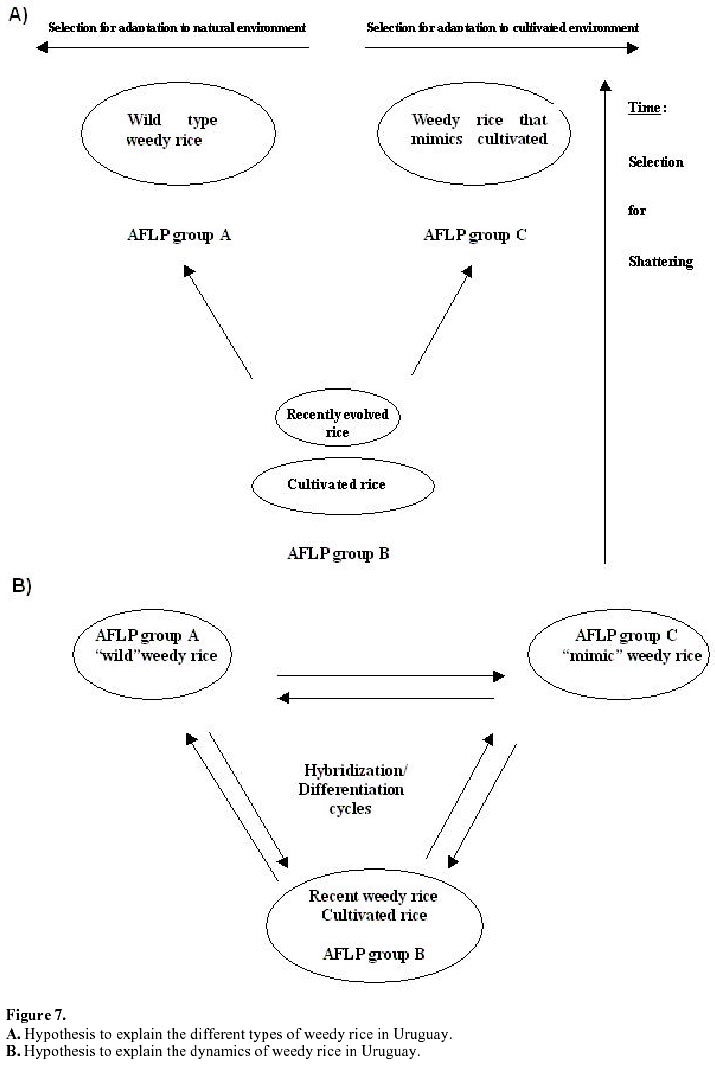
Each of the 10 primer combinations was able to generate an informative AFLP fingerprint across all 32 DNA samples including 26 weedy accessions and 6 cultivars (Table 1, Figure 2). Each primer pair produced between 12 and 44 distinguishable AFLPs bands with an average of (25.6) per primer pair (Table 3). About 130 unambiguous polymorphic AFLP bands were scored across the 32 samples, of which 127 bands were polymorphic among weedy material and 71 were polymorphic among cultivars. Associations among the accessions were investigated by UPGMA cluster analysis using NTSYS software (Figure 3). Samples formed three clusters based on genetic distances, two clusters (A and C) consisted of weedy samples only and one cluster (B) included all six cultivars analyzed and five weedy samples. Within cluster B there were two subgroups. One included two cultivars (INIA Tacuari, INIA Cuaro) and three weedy accessions (Tacuarembo-6, Unknown-2 and Rio Branco-5). The other included 4 cultivars (INIA Caraguata, Bluebelle, INIA Zapata, EEA 404) and two weedy accessions (Zapata-1, Unknown-3). Principal component analysis (PCA) based on genetic similarity (Figure 4) shows the relationships among accessions in two dimensional space. This shows that several weedy samples appear intermediate between groups, such as Tacuarembó-4, Vichadero-3 and unknown-3 and 4 (Figure 4). Associations among accessions were also analyzed by the Neighboring Joining Method (NJ) using AFLPDIST software to construct a matrix based on inferred nucleotide diversity, using a modified version of software developed by Innan et al. 1999 (Figure 5, Figure 6). Dendrograms with bootstrap values showing genetic relationships among accessions were constructed using Phylip software (Felsenstein, 1995) (Figure 5). The different analyses gave similar results. The samples in each of the AFLP groups were compared to geographic origin and morphological characteristics (Figure 1, Table 1). The weedy rice samples came from two major geographical regions of Uruguay, the east around Trent y Tres and about 80 km to the north around Tacuarembó department. While most samples in Group A are from the east (9 out of 12 samples) and most samples from the north were group C (4 out of 7) there was no clear trend. Based on morphological characteristics from AFLP group A, 75% (9 samples) have black hull and long or short awns. In group B about 90% of samples have straw hull and short or no awn. Group C consists only of samples with straw hull and no awn. Discussion The AFLP technique using silver staining to visualize bands has been successfully applied to the molecular characterization of Uruguayan weedy and cultivated rice. Two types of dendrograms were constructed to analyze genetic diversity between samples: one based on phenotypic data (NTSYS; Felsenstein, 1995) (Figure 3, Figure 4) and the other using phenotypic data to generate a genetic matrix that was subsequently analysed (AFLPDIST; Innan et al. 1999) (Figure 5, Figure 6). Both dendrograms showed similar results. Three clusters, A, B and C, were differentiated by both sets of analyses. Clusters A and B consist of only weedy rice samples whereas cluster B consists of all the cultivars and some weedy rice samples. No clear trend was detected between geographical origin of samples and genetic diversity. This may reflect the rather small sample number for which location was known and the limited geographic area from which samples were collected. It seems from the results that in northern and eastern rice growing areas of Uruguay all three genetic groups detected by AFLP analysis can be found. There was a clear relationship between AFLP groups and morphological characters. Wild type traits, such as, black hull and long awn was found in most of the samples in group A, whereas traits found in cultivated rice were found in the weedy rice of group B and C. It appears that weedy rice in Uruguay has two strategies for survival. The one is to adapt to the natural environment and the other is to mimic the cultigen. A hypothesis to explain the evolution of weedy rice in Uruguay is presented in Figure 7a. This hypothesis suggests that at different times there has been selection for shattering and subsequently disruptive selection for either wild traits (Group A) or domestication traits (Group C). Recently evolved weedy types that still are genetically similar to present day cultivars are represented by weedy samples in group B. At the same time hybridization-differentiation cycles between weedy rice and cultivars results in intermediate types that were found in this study such as Tacuarembó-4, Vichadero-3, unknown 3 and 4 (Figure 7b) (Harlan, 1975). The results of this study suggest that the control of weedy rice that have evolved from rice should consider the following:
Acknowledgments This research was conducted while the author was a JICA fellow of the Japanese government in Crop Evolutionary Dynamics Laboratory, Genetic Resources 2, National Institute of Agrobiological Resources, Tsukuba, Japan. I express my thanks to Dr. Duncan A. Vaughan, head of the Crop Evolutionary Dynmics Laboratory for receiving me and kindly supervising my work; to staff of the Crop Evolutionary Dynamics Laboratory particularly Dr. Xin Wang Wang for technical advice and Dr. Koji Doi and Dr. Akito Kaga for their assistance in computer processing. I express my thanks to Dra. Marta Francis (Unit of Biotechnology, INIA Las Brujas, Uruguay) for supporting my fellowship; and to Dr. Gonzalo Zorrilla and Ing. Nestor Saldain (INIA Treinta y Tres, Uruguay) for providing access and information on morphological characteristics of the samples. I am also grateful to Dr. Jordi García (IRTA, Barcelona) for corrections and suggestions about my work. References
Supported by UNESCO / MIRCEN network © 2001 by Universidad Católica de Valparaíso -- Chile The following images related to this document are available:Photo images[ej01026f7.jpg] [ej01026f5.jpg] [ej01026f4.jpg] [ej01026t3.jpg] [ej01026f1.jpg] [ej01026f3.jpg] [ej01026t1.jpg] [ej01026f6.jpg] [ej01026f2.jpg] [ej01026t2.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}