 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Electronic Journal of Biotechnology, Vol. 4, No. 3, December, 2001 The effect of protective ingredients on the survival of immobilized cells of Streptococcus thermophilus to air and freeze-drying Claude P. Champagne*1 and Nancy J. Gardner2 1Food
Research and Development Center, Agriculture and Agri-food Canada, 3600 Casavant,
Saint-Hyacinthe, Qué, J2S 8E3 Canada Tel: 514-773-1105 Fax: 514-773-8461
E-mail: ChampagneC@em.agr.ca Financial support: Goverment Research Center from Department of Agriculture and Agri-Food Canada. Received July 12, 2001 Code Number: ej01027 Abstract Streptococcus thermophilus cultures were grown either on trehalose or lactose, immobilized in alginate beads, dipped in various protective solutions and dried by either convection air-drying (CAD) or freeze-drying (FD). Immobilized cultures dipped in the 0.1% peptone solution did not show good survival to CAD or FD, as mortality was over 99%. There was no significant difference in mortality levels, in both methods of drying, when lactose or trehalose were used as protective ingredients. The highest survival levels (50 to 98%) were with a whey-sucrose protective medium, but this was potentially related to a higher pH and solids of the solution. Mortality levels were higher in FD than CAD, and this did not appear to be related to the fact that FD cultures had lower residual moisture contents than those dried under CAD. Cells grown on lactose had slightly higher survival rates to drying than those obtained from CAD. Trehalose-positive and trehalose-negative cultures of S. thermophilus did not show different mortality patterns to CAD or FD.
Keywords: compatible solutes, lactic acid bacteria,
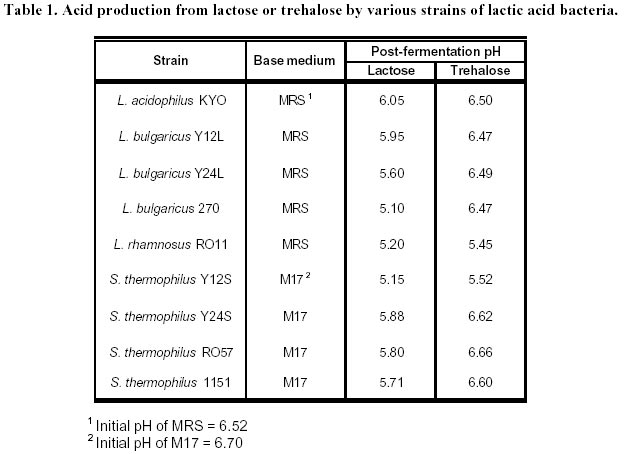
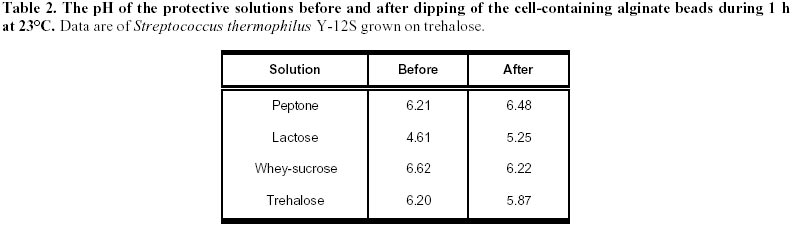
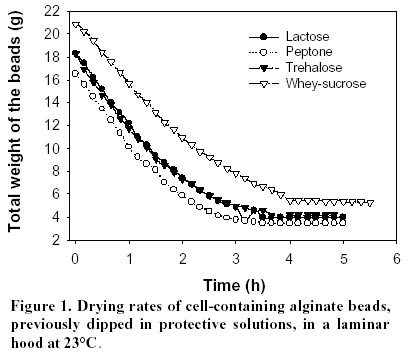
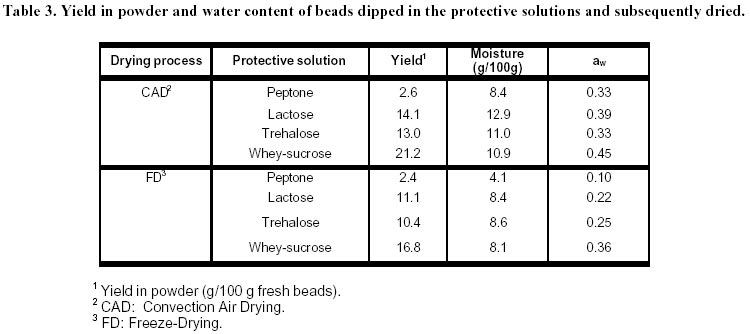
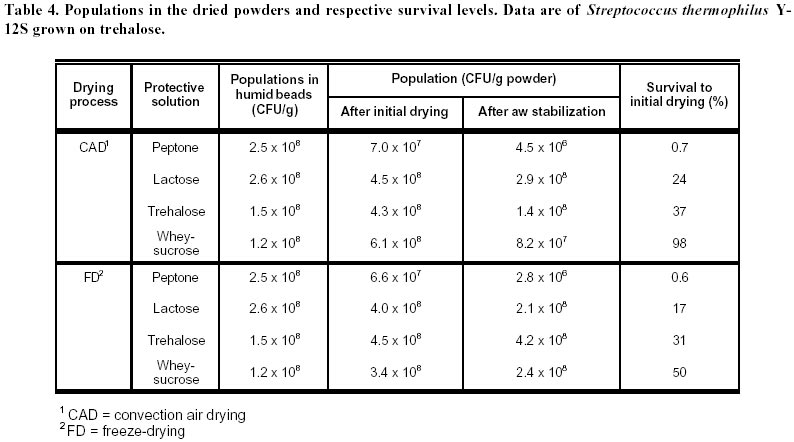
lyophilization. Article Lactic acid bacteria (LAB) are commonly used in the production of cheese, yoghurt, dry sausages, wine, sour-dough breads and sauerkraut. In many processes, a commercial culture is added directly to the food matrix, or in culture media for the production of starters. The commercial cultures available to the processors are marketed mostly in two forms: frozen and dried. The process used for the drying of food-related LAB is freeze-drying (FD), which is an expensive (Fellows, 1992) and slow method of drying. Bakers’ yeast, on the other hand, are dried by fluid-bed technology, and the application of such a convection air-drying (CAD) technology to LAB would be of interest to the producers of LAB starters, particularly in the emerging field of probiotic cultures. Many compounds have been shown to increase survival of LAB to freeze-drying (Kilara and Shahani, 1976; De Valdez et al. 1983), but few in CAD. The presence of compatible solutes in the medium, such as betaine or trehalose, has been shown to increase survival or microorganisms to air drying (Gadd et al. 1987; Leslie et al. 1995; Kets et al. 1996). The beneficial effect of these compounds, with respect to drying survival, seems related to their protective effect on proteins and membranes of the microorganisms (Wiemken, 1990; Colaço et al. 1992; Leslie et al. 1995). It was shown that survival to air drying was associated with an intracellular accumulation of the compatible solutes (Eleutherio et al. 1993; Kets et al. 1996). This accumulation is promoted by stressful growth conditions, particularly in response to the osmotic pressure of the medium (Hutkins et al. 1987; Molenaar et al. 1993; Glaasker et al. 1996). Saccharomyces cerevisiae is able to synthesize trehalose in response to the changes in osmotic pressures (D’Amore et al. 1991). However, synthesis of betaine or trehalose has not been demonstrated for lactic cultures, and these compatible solutes must be added to the growth medium and assimilated to be effective (Kets et al. 1996). The important literature involving the beneficial effect of trehalose to CAD drying incited us to evaluate this product in protective solutions. Although many LAB metabolize trehalose (Dellaglio et al. 1994) there are no reports on the intracellular accumulation of trehalose by the lactic cultures. However, it was shown in Saccharomyces cerevisiae that the presence of a trehalose carrier enhances the protective effect of trehalose added in the cell’s exterior environment (Eleutherio et al. 1993). Therefore, it was our hypothesis that the capacity of LAB to absorb trehalose would enhance the protective effect of this disaccharide to CAD of lactic cultures. The aim of this study was to evaluate the effect of various compounds in the growth medium as well as in the protective solutions on the survival of Streptococcus thermophilus cultures to CAD and FD. Materials and Methods Microorganisms. The list of strains screened in this study is provided in Table 1. Cultures were maintained in 100 mL of 12% (w/w) reconstituted non-fat dry milk (sterilized at 112ºC, 10 min). One transfer per week was carried out by inoculating milk with 2% of the strain and incubating at 37ºC for 16 h. Cultures were screened according to their ability to grow and to ferment trehalose. To do so, M17 or MRS broth in which trehalose was added as the sole carbon source were inoculated with 0.1% of the appropriate strain and incubated at 37ºC for 16 h. The M17 and MRS broths had the same formulation as the Difco (Detroit, USA) commercial products for all ingredients other than carbohydrates. With respect to carbohydrates, either lactose or trehalose were added in the media at 6 g/L. The pH was recorded before and after incubation. Immobilization. Streptococcus thermophilus cultures were prepared by inoculating M17 broths containing 0.5% trehalose or lactose with 0.1% of a milk-grown culture and incubating at 37ºC for 16 h. The culture was then centrifuged for 10 min at 6000 G using a Beckman J2-21 centrifuge. Cells were resuspended in 0.1% peptone (Difco, Detroit) to a concentration factor of 10X. The suspension was then added to an equivalent volume of 2% sodium alginate (BDH, Montreal) solution. The alginate-cell suspension was added dropwise using a 18G1½ needle (Becton-Dickinson, Rutherford USA), to an agitated solution (70 rpm) of CaCl2 0.1M. The beads were kept 1 h in the solution for gel strengthening before rinsing with 0.1% peptone. Drying. Beads (15 g) were added to 100 mL of the following solutions that were previously sterilized (121ºC, 15 min): 0.1% peptone (Difco), 14% whey (Saputo, St-Hyacinthe) supplemented with 6% sucrose (Lantic, Montreal), 10% lactose (Difco), 10% trehalose (Ajinomoto, Japan). The pH of all solutions was adjusted at 6.5 prior to autoclaving. To allow diffusion of the solutes, the beads were kept in the protective solutions for 1 h at 23ºC, recovered on sterile Buchner filters and sent to drying. For the air-drying treatment (CAD), beads previously dipped in the protective solutions were spread on a filter paper (Whatman 42, 15 cm, England) under a running laminar flow hood 16 h at 24ºC. For the FD process, beads were initially frozen at –70ºC and placed into a Calltech (FTS, Stone Ridge USA) freeze-dryer for 36 h with a drying temperature of 23ºC. The dried beads were weighed and analysed for viable populations and humidity. After the initial drying by air or FD, samples were taken for the determination of moisture content as well as viable counts, and the remaining beads (1 to 10 g depending on sample) were placed in a dessicator containing an over-saturated solution of LiCl2. The LiCl2 solution was added to generate an environment of 0.1 in available water (aw). The beads were left 5 days at 4ºC in the dessicator and viability as well a moisture were recorded. Analyses. For microbiological analyses, beads were placed in a 1% sodium citrate solution (100 mL) for 30 min. The suspension was then homogenized 1 min with a tissue homogenizer Omni 2000 (Omni International, Gainsville) set at 25000 rpm. Samples were subsequently diluted in sterile peptone 0.1% solutions. Enumeration of the streptococci was performed on M17 agar (Oxoid). Plates were incubated at 37ºC for 48ºh. For moisture content, beads were desorbed in methanol 24 h (Champagne et al. 1996) and water content was determined with the Karl Fisher method using the Mettler DL18 Karl Fisher Titrator (Zurich). The water activities (aw) were determined by placing samples at 23ºC in Rotronic Hydroskopt DT unit (Huntington, NY) which provided direct readings. Saturated solutions of lithium chloride (aw = 0.11) and magnesium chloride (aw = 0.33) were used to verify the accuracy of the unit. Results and Discussion Selection of the cultures It was demonstrated that the protective effect of trehalose to CAD drying was increased if this carbohydrate was found both on the internal and external sides of the cytoplasmic membrane (Eleutherio et al. 1993). Thus, the selection of a trehalose-positive strain was initiated in the hope that, in addition to the culture’s ability to metabolize trehalose, this strain would accumulate trehalose internally and show better survival to CAD. The capacity of various lactic cultures to produce acid from trehalose is found in Table 1. All strains fermented lactose and generated a significant drop in pH, but variations were noted with respect to trehalose fermentation. In the lactobacilli, L. rhamnosus RO11 seemed the best suited to assimilate trehalose. The strains used in this study thus showed the typical patterns of carbohydrate assimilation for their designated species (Dellaglio et al. 1994). Most of the Streptococcus thermophilus cultures also showed the typical trehalose-negative pattern (Dellaglio et al. 1994) except for strain Y12S (Table 1). In order to study strains of the same species, it was thus decided to use the strains Y12S and Y24S of S. thermophilus. Beads in the protective solution After recovery from the CaCl2, the beads were dipped into protective solutions prior to drying, and the pH of these solutions, before and after dipping of the alginate beads, are found in Table 2. Even though S. thermophilus is a thermophilic culture, there was acidification in the trehalose and whey-sucrose solutions during the 1 h incubation at 23ºC. The lowest post-incubation pH was in the lactose solution, but this is partially related to its low initial pH. The raise in pH is presumably related to the peptone/CaCl2 content of the beads, which could have neutralised the small quantity of acid of the unbuffered lactose solution. The dried products obtained Saccharomyces cerevisiae can be concentrated on vacuum drums supporting cellulose or starch filters to obtain particles of approximately 28% solids that can be subjected to fluid-bed drying. This procedure has not been applied to LAB and other means of providing a solid form to LAB concentrated cultures must be found. Immobilization in alginate beads was thus carried out for this purpose. Although CAD was used in this preliminary study, immobilization in alginate beads would enable fluid-bed drying of the cultures. The rate of drying in the CAD method is seen in Figure 1. In the conditions used, drying was complete after 4 h at 24ºC. The protective media in which the beads were dipped had different solute concentrations, from 0.1% for peptone and 20% with whey-sucrose. Thus, for a given initial amount of beads, the greater the solids content of the protective solutions, the greater the amount of powder was obtained following drying (Table 3). Additionally, the air cabinet-dried powders had higher humidity contents than the freeze-dried products, which resulted in higher aw readings. This must be kept in mind in examining the viability data. Indeed, the extent to which a product is dried will influence the survival rate of lactic cultures (Clementi and Rossi, 1984; De Valdez et al. 1985). Freeze-dried powders of LAB generally have between 1 and 4% residual moisture, but the FD products obtained in this study contained between 4.1 and 8.6% moisture (Table 3). This suggests that the drying period could have been extended, but the presence of alginate in the product could also account for the higher moisture levels. Indeed, it is well known that sugar-based compounds have different abilities in binding water (Fennema, 1996). Thus, for a given aw value, which reflects the level of free water, powders having compounds that bind more effectively water will have higher total moisture levels. This is because the powders have the same free water levels, but some have higher contents in bound water. In this respect, alginate has been shown to be more effective than disaccharides in binding water (Champagne et al. 1992). Since different protective solutions were used, the moisture composition of the dried beads varied accordingly. In order to determine if the level of drying had an influence on the survival rates, the beads were placed in a dessicator having a 10% relative humidity environment (aw of 0.1). Since all the dried beads had aw readings greater than 0.1 after the initial drying (Table 3), the products were placed at 4ºC for 5 days in the dessicator in order to try to have them all reach an aw level of 0.1. The goal of drying the products to identical aw levels was achieved, but readings (all between 0.27 and 0.34) showed much higher values than expected. Effect of the protective medium on survival to drying The population in the fresh beads was between 1.2 and 2.5 x 108 CFU/g (Table 4), which showed that dipping for 1 h in the protective solutions did not significantly change the viable population counts. However, important differences in viable populations in the dried products were obtained (Table 4). It must be noted that, as a consequence to the different amounts of powder obtained in each treatment, a comparison of the raw data of CFU/g of the powders obtained does not provide a good picture of the survival rates of the streptococci. For example, the biomass that was dried following the dipping of the 100 g of beads in the peptone solution was concentrated in only 2.5 g of powder, while the same population in presence of whey-sucrose was found in 16 to 21 g of powder (Table 3). Calculations were thus made so that the results presented would reflect the survival levels to the treatments with the following equation:
As had been found previously (Champagne et al. 1992), entrapment of cells in alginate alone does not offer them good protection to drying, since a drop of more than 99% in the viable population was observed when no protective solution was used. The protective medium in which the beads were dipped prior to drying had a significant effect on their survival. Peptone, at least at this low concentration, did not provide a good protection to drying. There was no significant difference between lactose and trehalose survival data. The whey-sucrose medium enabled the highest survival levels. However, it is not clear to which extent this is related to the medium ingredients and the pH of the medium. Having an appropriate pH does not, alone, guarantee a high survival level, as the data on the peptone medium shows. However, the whey-sucrose medium had a higher pH that the lactose and trehalose solutions (Table 2) and it has been shown that freezing LAB in acid conditions is detrimental to survival (Modler and Villa-Garcia, 1993). These results suggest that it is desirable to prepare protective solutions that not only contain compatible solutes, but also buffering compounds to adjust the pH. Whey contains phosphates and citrate that would provide this buffering capacity. A milk-sucrose-ascorbic acid medium is used commercially as a protective solution for freeze-drying of lactic cultures (Champagne et al. 1991). However, milk’s casein micelles are large and do not diffuse into alginate gels, and immobilized cells do not have access to these proteins (Passos et al. 1993). Consequently, when alginate beads are dipped into milk-based protective solutions, only the whey components can diffuse into the beads. This is supported by the finding that the whey-sucrose medium had been shown to be as effective in protecting alginate-entrapped cells to FD than commercial milk-based formulas (Champagne et al. 1996). Thus, the whey-based medium should reflect what could be expected in commercial formulations. Results show that this formulation gave the highest survival rates. This experimental plan does not enable us to determine why the whey-sucrose medium gave higher survival rates than the lactose/trehalose solutions, because of different solids levels, of the presence of two carbohydrates as well as of the whey solids. The whey-sucrose treatment is useful, nevertheless, in showing that using sugars alone, although providing a significant protection, are not as effective as are complex formulations typically used by industry. Effect of the strain and the type of sugar in the growth medium Strains Y12S and Y24S did not show different mortality patterns to drying (data not shown). As a whole, the effect of the protective medium was the most important on survival of the streptococci to CAD or freeze-drying. It must be emphasized that the effect of protective medium could be partially related to different pH values after dipping (Table 2) and that further studies are needed to determine the exact contribution of pH and medium ingredients to the effect on survival. Nevertheless, it was examined if the substrates used for the production of the trehalose-positive Y12S cells would influence the survival to drying. On the average, cells grown on lactose showed 44% less mortality to drying than cells cultured on trehalose (data not shown). However, the data suggests that this is more related to results with FD. It was hoped that producing the cells on trehalose would favour absorbtion of this compatible solute by the cells when they were dipped in the protective solutions. No determination of internal concentrations of trehalose or lactose were carried out, and it remains to be determined if indeed the growth medium influenced the physiology of the cells during the subsequent 1 h incubation of the beads in the protective solutions. The beneficial effect of trehalose in air drying of cells is well documented, but, unexpectedly, trehalose did not demonstrate a superior protective effect to CAD than lactose or sucrose. Two reasons might explain this. First, trehalose is not the only carbohydrate to act as a compatible solute (Csonka and Hanson, 1991) and it is possible that lactose and sucrose have equally protective effects for S. thermophilus Y12S. Secondly, the accumulation of compatible solutes is enhanced by osmotic stress (Hutkins et al. 1987; Molenaar et al. 1993). The experimental conditions of this study did not generate an osmotic stress, and thus may not have promoted the accumulation of the compatible solutes. More study is therefore needed on the possible interaction between carbohydrate growth substrate, osmotic stress during growth and internal accumulation of compatible solutes on survival levels of LAB to CAD. Effect of drying method With both strains, mortality levels were higher in FD, and paired t tests using the data of Table 4 showed this difference to be significant. It was our concern that this was related to the fact that the FD powders contained less water and had thus been submitted to a more extensive drying treatment. The incubation in the dessicator, carried out to stabilize aw values, resulted in lower viable counts in the dried powders, but the populations were still higher in the samples dried by CAD (Table 4). This suggests that the higher survival levels obtained in the CAD treatments were not related to the slightly higher residual moisture values. Concluding Remarks The high survival rates of S. thermophilus to CAD suggest that fluid-bed drying, used commercially for Saccharomyces cerevisiae, could successfully be applied to some lactic cultures. However, fluid-bed drying could only be applied to particulate concentrated cultures. Immobilization in alginate gels is instrumental in providing the solid support that is required for fluid bed drying. The advantage of alginate as a support is that it can easily be used for the production of concentrated suspensions of lactic cultures (Champagne et al. 1992; Champagne et al. 1993), and that the solid matrix can be dissolved upon rehydration, with citrates or phosphates, if the industrial application requires the use of the culture in a liquid form. There did not seem to be a difference between trehalose-positive and trehalose-negative strains with respect to survival to drying in general. Thus the capacity of LAB to metabolize trehalose does not seem to confer greater resistance to drying, but this needs to be examined more closely. In particular, strains could be selected not only for their capacity to absorb trehalose, but also for their ability to accumulate this carbohydrate, as do some yeasts. Although this study shows that CAD can be better than FD with respect to survival to the drying process itself, the stability to storage remains to be examined. The higher moisture content of the CAD products could translate into reduced stability during storage. This aspect will be crucial for the commercial exploitation of the technology, and studies are currently under way to examine this feature of the products obtained. Acknowledgments The technical support of R. Gera and L. Charlebois is gratefully acknowledged. References
Supported by UNESCO / MIRCEN network © 2001 by Universidad Católica de Valparaíso -- Chile The following images related to this document are available:Photo images[ej01027t3.jpg] [ej01027t1.jpg] [ej01027t4.jpg] [ej01027t2.jpg] [ej01027f1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}