 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Electronic Journal of Biotechnology, Vol. 4, No. 3, December, 2001 Molecular dynamics simulations of active site mutants of rat liver arginase Mauricio Canales*1, Linda Westermeyer2 and Nelson Carvajal3 1Laboratorio
de Biofísica, Departamento de Biología Molecular, Facultad de Ciencias
Biológicas, Universidad de Concepción, Casilla 152-C Correo 3, Concepción,
Chile Tel: 56 41 203822 Fax: 56 41 239687 E-mail: mcanale@udec.cl Financial support: Project 4533 from Universidad de Concepción, Chile to M.C.A. Received
July 12, 2001 Code Number: ej01028 Abstract By
using molecular dynamics (MD) simulations and crystallographic data for rat liver
arginase, the substrate positions in the active sites of native and mutant forms
of the enzyme, were compared and correlated with known kinetic consequences of
mutations. The mutants compared were His 141
Keywords: arginase, arginine, molecular dynamics,
optimization. Article Arginase catalyzes the hydrolysis of arginine to urea and ornithine. The enzyme is highly distributed in living organisms, in which it serves several functions (Aminlari and Vaseghi, 1992; Carvajal et al. 1995), including a role in the regulation of nitric oxide signaling (Chang et al. 1998). The crystal structure of rat liver arginase (Kanyo et al. 1996) shows a 15 Å deep active-site cleft in each monomer of the homotrimer, with two Mn2+ ions at the base of this cleft. Aspartate and histidine residues coordinate the metal ions and mutation of these residues has been shown to cause a lowering or loss of catalytic activity of the enzyme (Cavalli et al. 1994; Perozich et al. 1998; Carvajal et al. 1999). The
mutation His 141 For a better understanding of the kinetic consequences of replacement of His141 in arginase, we have now used molecular dynamics to compare substrate and inhibitor positions in the active sites of native and mutant forms of the enzyme. Materials and Methods Structures used and data management. The crystal structure of rat liver arginase was used as the basis of this study. As rat liver arginase is a homotrimer and it is known that each monomer is active, all of the studies were done only with the chain A of 1RLA (Kanyo et al. 1996). The identical B and C chains along with the water molecules that had been crystallized with them were removed. Water molecules in the active site, that obstructed the optimization of arginine but which were not coordinated with the metal ions, were also removed. The substrate arginine and the inhibitor lysine were inserted in the enzyme using the Insight II program (Biosym, 1995). The structure of arginine was inserted in an extended configuration with the guanidine group close to the metal ions and the main chain in the opposite direction. The structure used for arginine was that generated by the HyperChem database (Hypercube, 1992). Coordinates for lysine were those of arginine in the active site, changing only the atoms necessary to make arginine into lysine. Insertions were accomplished by saving the monomer coordinates in PDB (protein database) format and then transferring them to the Insight II program. This program was used at this point because of the ease it provides for manually localizing the substrate in the active site. After making the insertion the enzyme-substrate complex was returned to HyperChem and saved as Hin format used by HyperChem. Optimizations were done with AMBER 4.1 force field using a cutoff of 8.0 Å. Residues were modeled as charged forms in accordance to the pH of enzyme activity and the dielectric constant was assumed to be 1.0. The conjugate gradient algorithm from Polak-Ribiere was used with 200 iterations or 0.01 kcal/(Å mol)as termination condition. Substrate and inhibitor positions. Once inserted in the active site, the arginine was optimized to find the position of minimum energy for the substrate. In this process only arginine was allowed to move. The residues around the active site were held in position and the result of their forces on the substrate was studied. Since there was no charge available for Mn2+ at the time of this work, the metal ion was represented by the mass of Mn and its approximated van der Waals radius in the MD calculations. The same procedure was followed for lysine. Molecular Dynamics. After obtaining the optimized structure for arginine in the active site, MD was used to find the most stable average substrate position. Collection of data was done only after systems were equilibrated and total energy remained constant. Two series of MD simulations were carried out in order to fix the atomic coordinates that would be used as the control for the substrate and to find the optimum times and temperatures to be used. These conditions remained constant in the rest of the simulations and constitute the basis for comparison. One simulation was done for 20 ps at 300 K and the other for the same time period but at 500 K. Each simulation was done in two 10 ps parts. In each case the heating and cooling times were 0.1 ps and the step size was 0.001 ps. Data was collected every 5 time steps. Then, the snapshot files (data collected every 25 steps), were used to find the average positions. Since the two sets of simulations yield the same atomic coordinates for the substrate, the MD using 500 K was chosen and used to compare the atomic coordinates of the substrate or inhibitor in the wild type and mutants enzymes. The arginine was fixed in the active site using coordinates of the average position and then optimized again. This was the structure that was used as the basis of all comparisons. To find the average atomic coordinates for arginine in the MD simulation, the coordinates from the snapshot file of MD were converted to a text file using a script file, was used to run a replay of the MD and write the coordinates to a text file. For the analysis of the data an MS Access database was created. This same process was repeated to find the average displacement of the substrate or inhibitor from the atomic coordinates established for the control. MD
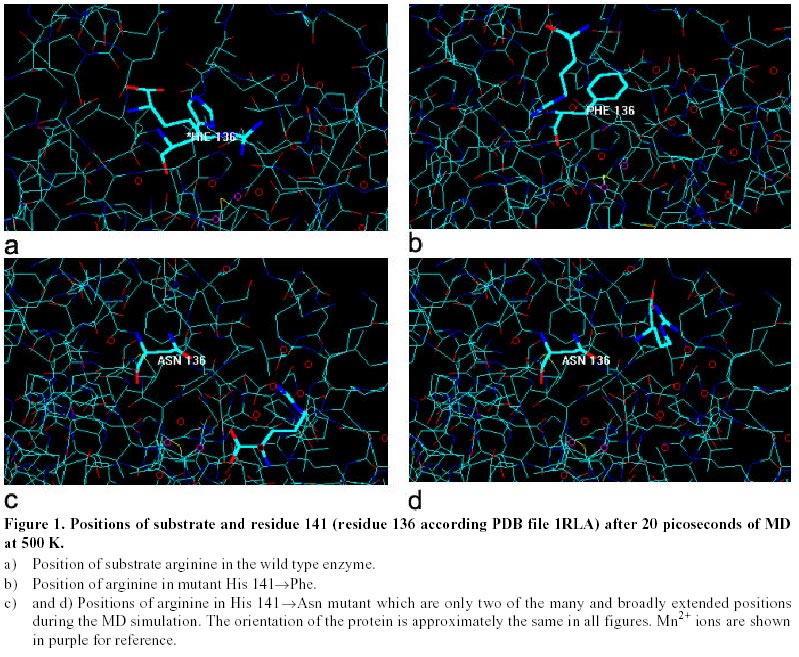
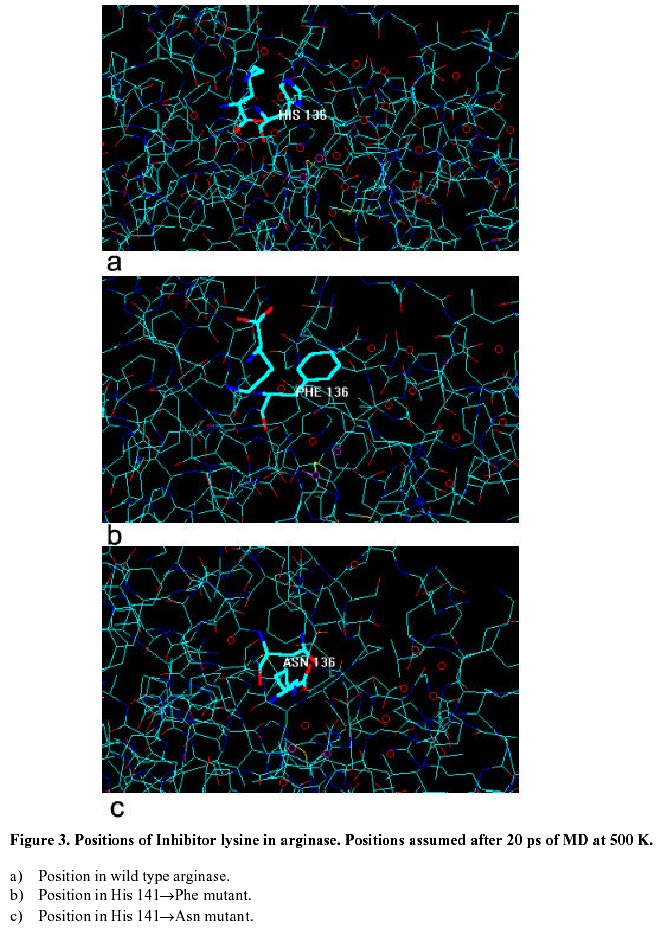
Simulation of mutated arginase. Mutations examined were His 141 Results and Discussion To successfully prepare a control position for the substrate, it was necessary to remove the crystallized water molecules that were at the active site, while retaining the water molecule involved in the coordination of the metals ions. It was assumed that the exclusion of these crystallized water molecules from the active site occurs spontaneously in the active enzyme. The two MD series at 300 K and 500 K yielded the same atomic coordinates for the arginine substrate. This was interpreted as confirmation of the probable position of the arginine in the active site during catalysis. To further confirm this position, the resulting structure was compared to that produced by the optimization of the substrate in Bacillus caldovelox (Bewley et al. 1999) followed by MD. There was close agreement between the two complexes. When the two structures were superimposed the guanidine group and the amino and carboxyl groups occupied nearly the same position. Since the MD series at 300 K and 500 K gave the same results, the model that resulted from the 500 K process was chosen as the basis for further studies. Figure 1a indicates the position that the substrate assumes in the active site during the MD. The substrate orients with the guanidinium group toward the metals with a distance of approximately 4.5 Å. The carbon chain of the arginine is approximately 4 Å from His141. The carboxyl group is about 3.6 Å from Asp 181 and 6.2 Å from Gly18. After optimization, the substrate was seen to have taken a position slightly out of the active site of the H141F mutant enzyme, Figure 1b. During MD the substrate maintained its attraction for the active site and repeatedly moved out of and then backs into the active site, while maintaining its extended structure. However, the substrate never completely returned to the optimized position that it had before the mutation. A much more drastic change was produced by mutation of His141 by asparagine. The substrate never assumed a fixed position and continuously travelled around the H141N enzyme molecule, Figure 1c and Figure 1d. Mutant
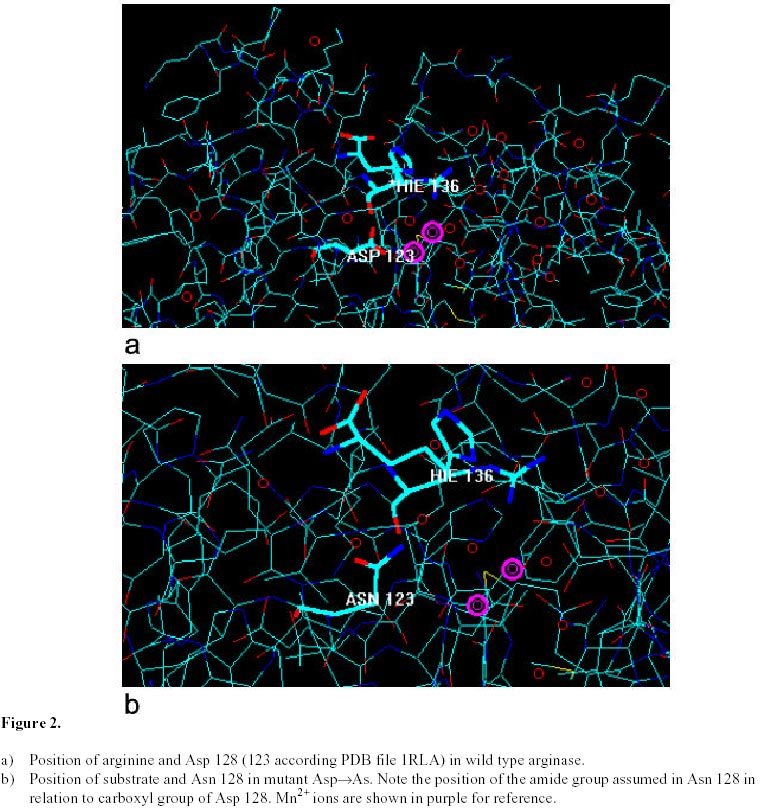
Asp128 In
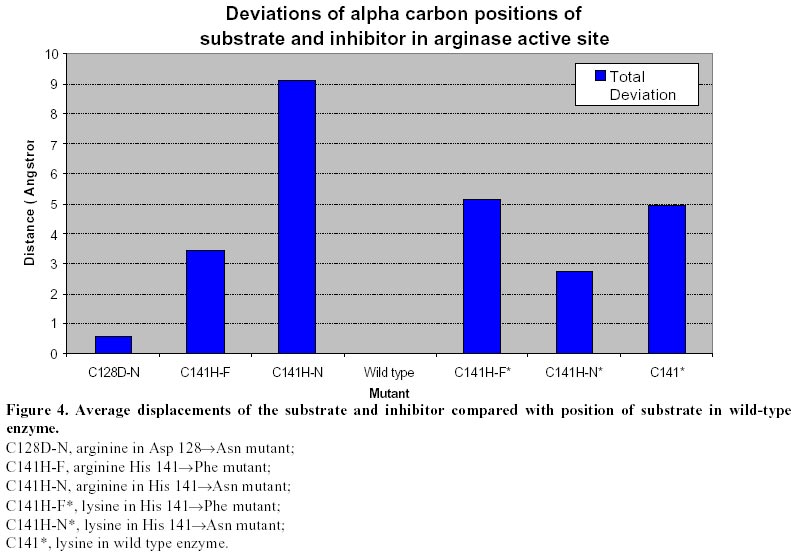
conclusion, theoretical MD simulations revealed significant differences in the
average displacements of the substrate molecule in the active site of wild-type
and mutant enzymes. Interestingly, the more drastic differences were noted for
the H141N mutant. This is interesting if one considers the kinetic consequences
of replacement of His141 by phenylalanine or asparagine. Although both H141F and
H141N exhibit about 10% of wild-type activity, only the His141 Theoretical MD simulations are of necessity very short and, for this reason; it is impossible to see all of the possible positions that may be occupied by a molecule. In any case, some conclusions may be drawn about the position of the substrate in the enzyme molecule. On the other hand, the usefulness of the comparative approach used here and the validity of the conclusion that mutation of His141 by asparagine is significantly more drastic than a change by phenylalanine is reinforced by the observation of nearly the same position for the substrate in wild-type and D128N enzymes. Asp-128 is known to interact with the metal ion and not with the substrate in arginase (Kanyo et al. 1996; Perozich et al. 1998). Acknowledgments We thank the Graduate School from Universidad de Concepcion for the support to L.W. during her Master studies. References
Supported by UNESCO / MIRCEN network © 2001 by Universidad Católica de Valparaíso -- Chile The following images related to this document are available:Photo images[ej01028f3.jpg] [ej01028f1.jpg] [ej01028f2.jpg] [ej01028f4.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}