 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Electronic Journal of Biotechnology, Vol. 4, No. 3, December, 2001 Trichoderma aureoviride 7-121, a mutant with enhanced production of lytic enzymes: its potential use in waste cellulose degradation and/or biocontrol Mercedes Zaldívar*1, Juan Carlos Velásquez#2, Inés Contreras3 and Luz María Pérez§4 1Departamento
de Bioquímica y Biología Molecular, Facultad de Ciencias Químicas
y Farmacéuticas, Universidad de Chile, PO Box 174, Santiago 22, Chile Tel:
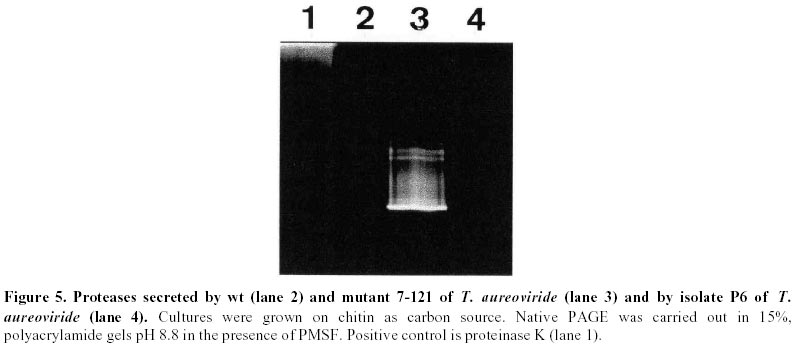
56 2 6781658 Fax: 56 2 2227900 E-mail: mzaldiva@uchile.cl Financial support: IFS Grant E/1159 (M.Z.) and Fondecyt 1970532 (L.M.P.). Received July 19, 2001 Code Number: ej01029 Abstract A mutant of the native fungus Trichoderma aureoviride, 7-121, selected for its overproduction of extracellular cellulase and ß-glucosidase (cellobiase) was obtained. In shake flask cultures, production of endoglucanase, filter paper activity and cellobiase increased two to four- fold as compared with the wild type strain. The mutant strain is stable and grows rapidly in liquid as well as in solid culture media. Enzyme yields were best when pH was controlled so that it did not fall bellow pH 3.5. Cellobiase production by this mutant is particularly high (approximately 5 U/ml) as compared to other Trichoderma, strains, which makes it a suitable candidate for waste cellulose degradation. In addition, the mutant strain showed enhanced production of fungal cell wall degrading enzymes: chitinases, ß-1,3-glucanases and proteases. This improvement in extracellular enzyme production by the mutant T. aureoviride 7-121 suggests that it is a suitable strain to be used in biological control. Keywords: biological control, lytic enzymes, Trichoderma aureoviride. Article Members of the fungal genus Trichoderma have been extensively studied, particularly due to their ability to secrete cellulose degrading enzymes or to act as biocontrol agents. Most of the work has been carried out on strains of T. viride, T. reesei, T. harzianum. These strains have been extensively studied in their ability to produce extracellular cellulolytic enzymes, namely endoglucanases, exoglucanases and cellobiase, which act synergistically in the conversion of cellulose to glucose (Eveleigh, 1987). The strains have been mutagenized and genetically modified to obtain an organism capable of producing high levels of cellulases (Mandels and Andreotti, 1978; Nevalainen et al. 1980; Durand et al. 1988; Szengyel et al. 2000). The objective of these efforts has been to obtain high levels of cellulases in order to degrade waste cellulose, and its application in animal feed, pharmaceutical, textile and pulp and paper industry. However, despite the effort of many laboratories, no commercially efficient enzyme complex has been produced. The high cost of enzyme production limits the industrial use of the enzymes in the production of soluble sugars. In addition, the Trichoderma cellulase system is deficient in cellobiase, causing the accumulation of the disaccharide cellobiose, which produces repression and end product inhibition of the enzymes, both of which limit enzyme synthesis and activity. As a result, production of glucose from waste cellulose is still not commercially feasible. Furthermore, members of the genus Trichoderma, particularly T. harzianum, have also been reported as biocontrol agents (Lewis and Papavizas, 1991; Haran et al. 1996a; Haran et al. 1996b; Elad, 2000; Hermosa et al. 2000) due to its ability to successfully antagonize other fungi including plant pathogenic species. There are several mechanisms involved in Trichoderma antagonism namely antibiosis whereby the antagonic fungus shows production of antibiotics; competition for nutrients; and mycoparasitism whereby Trichoderma directly attacks the plant pathogen by excreting lytic enzymes such as chitinases, ß-1,3 glucanases and proteases (Haran et al. 1996a). Because the skeleton of filamentous fungi cell walls contains chitin, glucan and proteins, enzymes that hydrolyze these components have to be present in a successful antagonist in order to play a significant role in cell wall lysis of the pathogen (Lorito et al. 1994; Carsolio et al. 1999). Several distinct chitinolytic enzymes have been reported in T. harzianum (De la Cruz et al. 1992; Haran et al. 1996b). These include endochitinases, exochitinases and 1,4-ß-N-acetylglucosaminidases, which are induced during growth of T. harzianum in liquid medium containing chitin as carbon source. Another enzymatic system that is involved in cell wall degradation by an antagonistic organism is ß-glucan degrading enzymes. Two mechanisms of glucan degradation have been reported: exo- and endo-glucanases, both of which act synergistically in glucan degradation. ß-glucan degrading enzymes are classified according to the type of ß-glucosidic linkages: 1,4-ß-glucanases (including cellulases), 1,3-ß-glucanases, and 1,6-ß-glucanases (Pitson et al. 1993). Because 1,3-ß-glucan is a structural component of fungal cell walls, the production of extracellular 1,3-ß-glucanases has been reported as an important enzymatic activity in biocontrol microorganisms. In addition to chitin and glucans, filamentous fungi cell walls contain proteins. Thus, the production of proteases may play a role in antagonism (Sivan and Chet, 1989; Flores et al. 1997). Our laboratory has been interested in the production of enzymes by native Trichoderma strains, in particular by T. aureoviride, to obtain high levels of extracellular cellulases and cell wall-degrading enzymes for biological control. The present work characterizes a mutant strain of T. aureoviride, 7-121, selected by its ability to secrete cellulases and ß-glucosidase, and studies the production of extracellular cell wall degrading by the wild type and mutant strains. Materials and Methods Fungal strains. Trichoderma aureoviride strains were isolated from soil samples and from sooty molds growing on Citrus trees (Musalem et al. 1984; Pérez et al. 1991). They were selected for their ability to grow on cellulose as sole carbon source. Isolated strains were grown on Potato Dextrose Agar (Difco) plates for 5 to 7 days at 28ºC. Cultures were stored at 4ºC and transferred weekly. Induction of mutants. Conidia from one-week-old PDA plates were suspended in 9 g/L sterile NaCl solution. A suspension containing 5x108 conidia/mL was treated with 200 µg/mL N-methyl-N-nitro-N-nitrosoguanidine (NG, Sigma) for 30 min at 37ºC in a water bath shaker. At this dose, 90% mortality is achieved. Surviving conidia were cloned on PDA plates with the addition of 0.1% Triton X-100 and 4 g/L L-sorbose as colony restrictors (Morikawa et al. 1985) Plates were incubated at 28ºC for four days. Isolated colonies were replicated onto the plate-screening medium. Plate screening medium. This medium contained Mandel’s mineral salts solution (Mandels et al. 1974) with the addition of 17.5 g agar/L, 5.0 to 10 g phosphoric acid-swollen cellulose/L (Tansey, 1971), 0.1% (v/v) Triton X-100 and 4 g L-sorbose/L. Plates were incubated at 28ºC for 3 days followed by 18 h at 50ºC. Hypercellulolytic mutants were selected on the basis of the diameter of the hydrolysis zone surrounding the colonies. Production of extracellular enzymes in shake flasks. The production of extracellular enzymes was induced in submerged cultures of the wild type and mutant strains. Cultures were carried out in Mandels Salts solution with the addition of 5 or 7.5 g/L of the appropriate carbon source. Conical flasks containing 200 mL of Mandels medium were inoculated with 5x106 conidia. Cultures were incubated in an orbital shaker incubator for 14 days at 28ºC. At two days intervals, culture aliquots were centrifuged at 12.000xg to remove solids. The supernatants were assayed for enzymatic activities. Carbon source. For the production of chitinases, liquid cultures were carried out using white chitin as sole carbon source. ß-1,3-glucanases was induced by the addition of yeast cell wall glucans (Manners and Wilson, 1973). ß-1,4 glucanases, ß-glucosidase and filter paper activity (overall cellulolytic activity) was induced by the addition of microcrystalline cellulose (Sigmacell type 50) in the culture media. In addition, fungal cell walls were used as carbon source. They were extracted according to the method of Sivan and Chet (1989), from Botrytis cinerea, Alternaria alternata and Fusarium oxysporum. Enzymatic Assay procedures. ß-1,4-Endoglucanase activity: This was assayed by the method proposed by Novo Industry, using carboxymethylcellulose (substitution degree 0.7, Sigma) as substrate. Reaction mixtures containing 50mM sodium acetate pH 5.0, 4 g CMC/L and culture supernatant, were incubated for 10 min at 50ºC. The formation of reducing sugars was measured with potassium ferricyanide in alkaline conditions. Filter paper activity (FPA): This method estimates overall cellulolytic activity. It was assayed by a modification of the method of Mandels (1974), incubating 0.5 mL of a dilution of the culture supernatant with 50 mg of filter paper (Whatman Nº 1) in 0.5 mL of 0.04 M acetate buffer (pH 5.0) at 50ºC for 60 min. The liberated reducing sugars were assayed by the Somogyi-Nelson method (Nelson, 1944). ß- glucosidase activity: 0.1 mL of the culture supernatant was incubated with 0.5 mL of 0.05 M acetate buffer (pH 5.0) containing 2.5 mg cellobiose. After incubation at 50ºC for 10 min the glucose released was measured by the glucose oxidase peroxidase method, (Sigma). Enzymatic activity of the enzymes of the cellulase complex were expressed as International Units/mL defined as the amount of enzyme which releases one micromole of reducing sugars expressed as glucose per minute. Non enzymatic controls were performed using boiled enzymes and were subtracted from the corresponding enzymatic values. Exochitinase activity: Culture supernatants were incubated with p-nitrophenyl-N-acetyl-glucosaminide (130 µg/µL) for 3 min at 30ºC. Reaction was stopped by the addition of 1 M NaOH. The released p-nitrophenol was measured spectrophotometrically at 410nm. The enzymatic activity is expressed as µM of p-nitrophenol released per minute. Blanks were carried out using boiled enzyme, and sterile culture medium. Endochitinase activity: Proteins in the culture supernatants were separated by polyacrylamide gel electrophoresis. Samples contained 20% sucrose and 0.01% (w/v) bromophenol blue (anodic PAGE) or 0.01% (w/v) methylene blue (catodic PAGE). Native PAGE were carried out according to the method described by Pan et al (1991). Stacking gels contained 4% acrylamide and running gels contained 15% acrylamide (acrylamide/bis-acrylamide 30:0.8). After electrophoresis at 30mA, the endochitinase activity was assayed in an agarose gel containing 0.04% glycolchitin in 0.1M sodium acetate pH 5.0. The glycolchitin agarose gel was place in contact with the acrylamide gel and incubated at 40ºC for 90 min. The agarose gel was then submerged for 5 min. in a freshly made solution containing 0.01% fluorescent brightener 28 in 0.5M Tris-HCl pH 8.9. The gel was then washed with distilled water, and incubated at room temperature in the dark for 2 hours, or alternatively at 4ºC overnight. Gels were observed with UV light. Endochitinase activity was visualized as dark bands against a clear background. Serratia marcescens chitinase was used as positive control. ß,1-3 glucanase: Proteins in the culture supernatant were separated by native PAGE as above. After electrophoresis the polyacrylamide gel was incubated with a solution of 1% laminarin in 50mM sodium acetate pH 5.0 at 37ºC for 45 min. The gel was then fixed in acetic acid: methanol: water (5:5:2) for 5 min, washed thoroughly with distilled water, and it was then incubated with 0.15% triphenyltetrazolium in 1M NaOH. The gels were heated in a microwave oven. ß,1-3 glucanase were visualized as red bands (Sivan and Chet, 1989). Protease activity: Protease activity was analyzed in the culture supernatants of the strains grown in cellulose, chitin, or glucan. Proteins were separated by native PAGE. Before casting the gels, 2% and 6% bovine hemoglobin was added to the stacking and running gels, respectively. Electrophoresis was carried out at 4ºC and 27mA. The gel was then incubated in a solution containing 20mM sodium succinate pH 4.0 and 10 mM 2- mercaptoethanol for 3 h at 40ºC. The gel was then stained overnight in a 0.025% Coomasie Blue R-250 solution in 25% isopropanol and 10% acetic acid. Protease activity was visualized as clear bands against a blue background. Proteinase K was used as positive control. Myceliar protein: biomass was measured as myceliar protein. Cultures were grown in 125 mL conical flasks containing 20 mL of medium. At the desired time intervals, the contents of a flask was centrifuged at 4,500 x g, the pellet was washed, resuspended in saline and homogenized. An aliquot of the homogenized suspension was extracted with one volume of 1N NaOH at 50ºC for 45 min and centrifuged. Proteins were measured in the supernatant by the method of Lowry, after precipitation with 20% TCA. Results
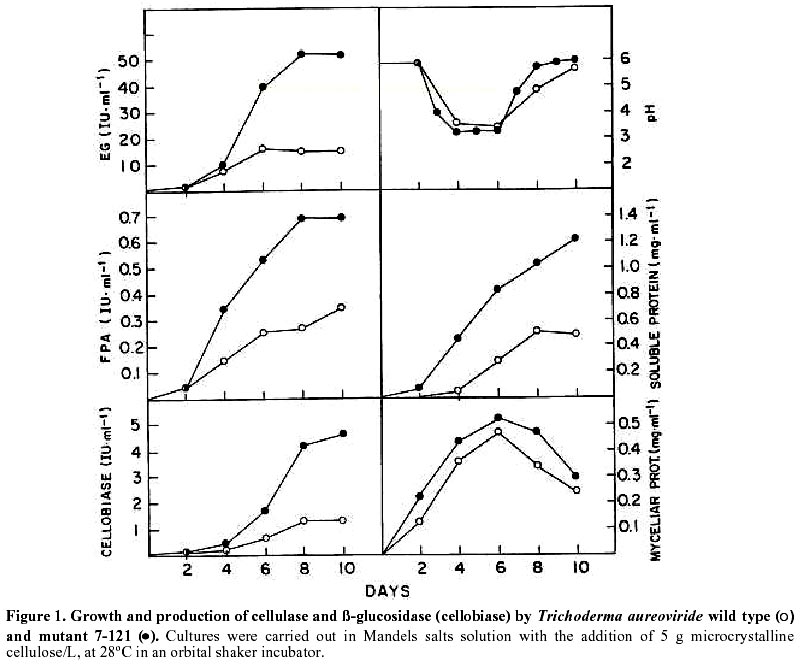
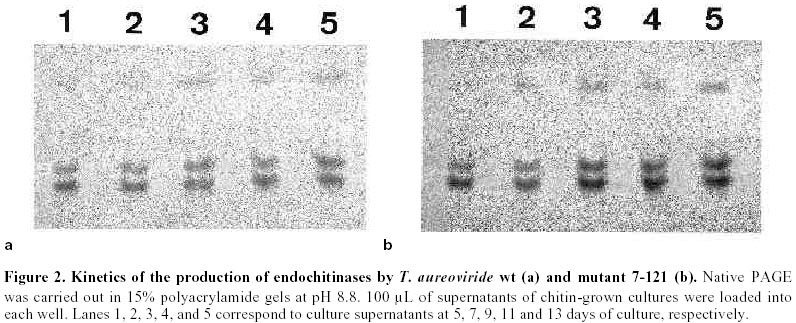
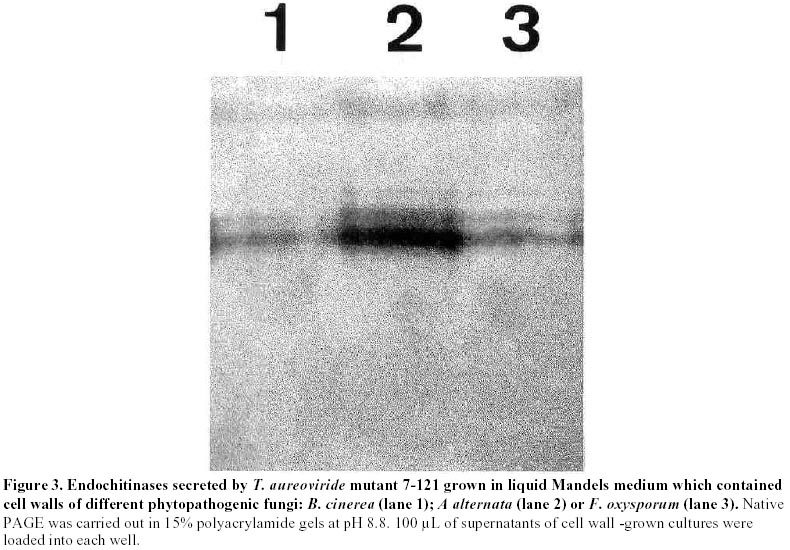
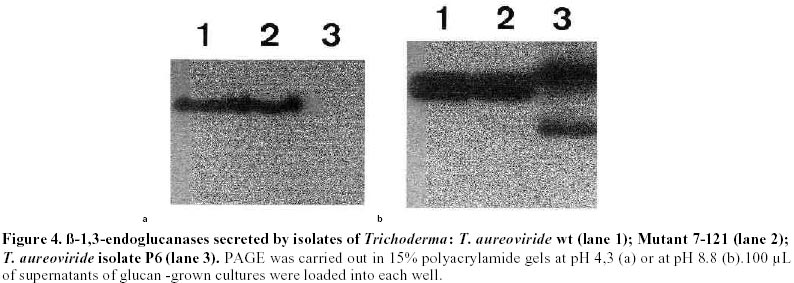
Mutagenesis of T. aureoviride A wild type strain, isolated by its ability to grow on cellulose as carbon source was subjected to successive mutagenic treatment with NG. After four mutagenic steps a mutant strain 7-121 was isolated. The selection of hyperproducing cellulase mutants was based on the diameter of the clearing zone surrounding the colony on the plate-screening medium. This screening medium was found to give fairly reliable indication of elevated cellulolytic activities. In the third and fourth mutagenic steps the concentration of cellulose was increased from 5 to 10 g/L. After each mutagenic treatment, enzyme production by the clones that showed larger clearing zones was assessed in shake flask cultures, and the most promising strain was subjected to the next mutagenic treatment. Enzyme production by T. aureoviride wild type strain and mutant 7-121 Figure 1 shows the production of cellulase and ß-glucosidase by mutant 7-121 as compared to the wild type strain. The mutant strain secreted approximately three times more ß-1,4-endoglucanase than the wild type, twice the FPA and roughly four times more ß-glucosidase. As shown in Figure 1, the rate at which the enzyme appeared in the culture medium was higher in the mutant 7-121. It should be noted that the levels of ß-glucosidase observed for both wild type (1.25 U/mL) and mutant strain (4.9 U/mL) are unusually high for a species of Trichoderma. No difference in the growth of both wild type and mutant strain was observed as assessed by measurement of myceliar protein. This result indicates that the enhancement of enzyme production by the mutant strain is not due to an increase in growth but to the enhancement in production and/or secretion of the enzyme Production of chitinases: Figure 2 shows the production of endochitinases by the wild type strain and its derived-mutant 7-121 induced by chitin. The kinetics of enzyme production by the wild type strain showed three bands of activity that appear at day 5 of culture (Figure 2a). Mutant 7-121 also produces three bands of extracellular endochitinases after day 5, although the intensity of the bands are clearly stronger (Figure 2b), suggesting that in the mutant strain, the enzyme secretion is enhanced. In order to test the induction of endochitinases by fungal cell walls, mutant 7-121 was grown on fungal cell wall extracts as sole carbon source. Three bands of endochitinase activities are observed as a result of growing the mutant using B. cinerea cell walls as substrate (Figure 3, lane 1), while in the presence of cell walls from A. alternata (Figure 3, lane 2) or from F. oxysporum (Figure 3, lane 3) four bands of enzyme activity are visualized. Best induction is achieved in the presence of A. alternata cell walls. The electrophoretic pattern of endochitinases is similar to that obtained in chitin (Figure 2b) suggesting that similar isoforms are induced by all substrates. Therefore, T. aureoviride mutant 7-121 can effectively induce chitinolytic enzymes in the presence of the plant pathogens mentioned above, indicating that this mutant is an interesting candidate to be used for biological control of these pathogens. Production of exochitinase: supernatants were assayed for exochitinase activity after 12 days of culture in the presence of chitin or fungal cell walls. Both wt and mutant strains produced low levels of this enzyme, even after 25 days of culture. The highest activity were 1.2 and 2.3 U/mL for wt and mutant strains, respectively. When grown on fungal cell walls, exochitinase production by the mutant strain reached similar levels as those obtained in chitin, namely 1.9 U/mL in A. alternata, 1.5 U/mL in F. oxysporum and 0.9 U/mL in B cinerea. These results indicate, that although exochitinase production by both strains is low, it is roughly the same in both purified chitin as well as in fungal cell wall. Production of ß-1,3 glucanase: Figure 4 shows the results of enzyme production by the wt and mutant strain (lanes 1 and 2, respectively). It is compared to enzyme production of a second isolate of T. aureoviride (lane 3). One band of ß-1,3-endoglucanase activity for wt and mutant 7-121 is observed when analysis was done in acidic conditions (Figure 4a, lanes 1 and 2). A second isolate of T. aureoviride shows two bands of activity (Figure 4a, lane 3). However, analysis under alkaline conditions (Figure 4b) shows that the only visible bands are those produced by the wt and its derived-mutant strain. Glucanase activity was also detected in cultures grown in chitin, in the absence on ß-glucans (data not shown). In this latter case, both strains produce two activity bands when analysis was performed under acidic conditions. Two bands of ß-1,3-glucanase are also observed when the wt and 7-121 strains are grown in fungal cell walls. In this case, best enzyme production is apparent in A. alternata cell walls (data not shown). These results suggest that the production of ß-glucanase by T. aureoviride is constitutive. Production of extracellular proteases: Mutant 7-121 grown in chitin shows several bands with protease activity (Figure 5, lane 3) as compared to its parental strain (Figure 5, lane 2) and another isolate of T. aureoviride (Figure 5, lane 4). The number and distribution of protease bands was not affected by the addition of PMSF, suggesting that these are not seryl proteases. The same bands were observed when the mutant strain was grown on cell wall extracts. It should be noted that although protease activity was clearly detected, it does not affect the detection of chitinase in the same supernatant, suggesting that chitinases from these fungi are not sensitive to proteases secreted by themselves. Discussion The enzymatic degradation of waste cellulose by fungal enzymes has been suggested as a feasible alternative for the conversion of lignocellulosics into fermentable sugars and fuel ethanol (Oksanen et al. 2000; Shin et al. 2000; Szengyel et al. 2000). However, the most widely studied enzymatic system, namely Trichoderma cellulases, has shown several disadvantages. Attempts to use these enzymes in the degradation of cellulosic wastes have not been successful for several reasons such as: low enzymatic yields, low specific activities, end product inhibition of the enzymes. One of the reasons which has been reported for low enzyme yields, is catabolite repression of enzyme biosynthesis due to cellobiose; Trichoderma are deficient in ß-glucosidase (cellobiase), therefore cellobiose accumulates and represses enzyme biosynthesis. Most of the cellulolytic strains of Trichoderma described in the literature are deficient in the production of ß-glucosidase (Durand et al. 1988). Mutant 7-121 however produces high levels of cellobiase, which can be further improved by controlled culture conditions. When urea was used as Nitrogen source, and pH levels remained within the 4 to 6 range, cellobiase yields were improved (data not shown). In addition, the mutant strain produces good levels of endoglucanase and FPA, which together with the exceptionally high levels of cellobiase, makes this mutant a good candidate for obtaining cellulases for lignocellulosic waste degradation. Because Trichoderma species have been reported as biocontrol agents (Grondona et al. 1997; Lorito et al. 1998; Baek et al. 1999; Carsolio et al. 1999) we further investigated the production of other extracellular enzymes by T. aureoviride wild type and mutant 7-121. One of the major mechanisms involved in the antagonistic activity of members of the genus Trichoderma against phytopathogenic fungi, is mycoparasitism (Haran et al. 1996a; Haran et al. 1996b; Lorito et al. 1998). In order to be able to degrade phytopathogen cell walls, the antagonistic Trichoderma induces the production of extracellular hydrolytic enzymes, responsible for the direct attack against the pathogen. These enzymes have been reported mainly in isolates of T. harzianum (Lorito et al. 1993; Lorito et al. 1994; Haran et al. 1996a; Haran et al. 1996b; Lorito et al.1996; Lorito et al. 1998). Cell wall degrading enzymes of different isolates of T. harzianum have been extensively studied; several isoforms of the enzymes have been reported. The genes involved in their biosynthesis have been identified and cloned, the protein structure has been deduced from the nucleotide sequence (García et al. 1994; Hayes et al. 1994). The enzymes have been purified and the effect of the purified protein has been assayed on spore germination and elongation. Considerable variation has been reported with respect to biocontrol activity and host range among the Trichoderma species analyzed. The production of the hydrolytic enzymes has been shown to be affected by culture conditions and by the host (De la Cruz et al. 1992; Lorito et al. 1994). Our laboratory decided to test as a biocontrol agent, a mutant strain of T. aureoviride, 7-121, which was originally isolated by its ability to overproduce cellulases. Mutant 7-121 was able to grow on both chitin and ß-glucans as sole carbon source. The production of extracellular chitinases and ß-glucanases was demonstrated. In both cases there is an apparent improvement in enzyme secretion as compared to the wild type strain. In addition the enzymes were also produced in the presence of phytopathogen cell walls as carbon source, suggesting that these substrate can also act as inducers of the synthesis of the lytic enzymes enzyme. This result is similar to that reported by Sivan and Chet (1989) in which T. harzianum produced high levels of chitinase and ß-1,3-glucanase when grown on R. solani mycelia. Proteases are also induced during growth in both chitin and ß-glucans. Electrophoretic patterns indicate that there are several proteolytic enzymes induced. However these enzymes have no apparent effect on either chitinases or ß-glucanases. Protease role in antagonism has been reported in experiments in which the susceptibility of F. oxysporum hyphae to chitinase and ß-glucosidase is increased after pretreating hyphae with protease (Sivan and Chet, 1989). Our results indicate that T. aureoviride, mutant 7-121, overproduces cellulases ß-glucosidase, and the extracellular enzymes responsible for antifungal activity, which suggests that this strain may be utilized as a biocontrol agent and/or a source of enzymes for cellulose degradation. In addition, this strain has proved to be very stable, showing little or no reversion, and grows rapidly in both liquid and solid media; formation of conidia is normal and its enhanced levels of cellulase production have remained unchanged for over a year. Acknowledgments The authors which to thank Dr. Eduardo Piontelli who identified the isolates of Trichoderma aureoviride. References
Supported by UNESCO / MIRCEN network © 2001 by Universidad Católica de Valparaíso -- Chile The following images related to this document are available:Photo images[ej01029f2.jpg] [ej01029f3.jpg] [ej01029f1.jpg] [ej01029f5.jpg] [ej01029f4.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}