 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Electronic Journal of Biotechnology, Vol. 5, No. 1, April, 2002 Isolation and partial purification of a metabolite from a mutant strain of Bacillus sp. with antibiotic activity against plant pathogenic agents Giuliano Bernal * #1, Andrés Illanes2 and Luigi Ciampi3 1Instituto
de Producción y Sanidad Vegetal, Facultad de Ciencias Agrarias, Universidad
Austral de Chile, Isla Teja, Chile E-mail: gbernal@canela.med.uchile.cl
Financial support: Grant 1951105 from Fondecyt. Received
November 15, 2001 Code Number: ej02009 Abstract The use of microorganisms for biological purposes has become an effective alternative to control plant pathogens. There are many examples of formulations using bacterial or fungal strains with biocontrol applications. Among them, members of the genus Bacillus are well known antibiotic producers. However, the increased capacity of antibiotic production obtained by direct mutagenesis of wild strains, has seldom been reported in the open literature. This research refers to the mutation of the A47 Bacillus strain, a plant pathogen antagonist, in order to obtain an improved strain with enhanced capacity to synthesize metabolites with antibiotic activity. The mutant strain M40 was obtained using the mutagenic agent acridine orange. The mutant strain showed a higher antagonistic activity than the wild type A47 against the plant pathogen Botritys cinerea (grey mould), Ralstonia solanacearum (bacterial wilt) and Erwinia carotovora var. carotovora (bacterial soft rot). The final objective was to isolate the antibiotic metabolite produced by the M40 strain and to determine its chemical and antibiotic properties. The results revealed the presence of an extracellular, thermostable and methanol-soluble metabolite that absorbed light at 212nm. These characteristics are similar to those described for cyclic antibiotic lipopeptides such as iturins. Keywords: Bacillus subtilis, biological control, iturin. Article The biological control of plant pathogens is of paramount importance nowadays, since the conventional use of chemical pesticides has been seriously questioned because of environmental and human health hazard (Spurrier, 1990; Mendgen et al. 1992).
Silva (1992), isolated a strain from the surface of healthy raspberry fruits that showed a very high antagonistic activity against several strains of Botrytis cinerea, an important post harvest plant pathogen of grapes, berries and other small fruits. This antagonistic isolate, designated as A47, that was active against Botrytis cinerea in vitro and in vivo, showed also similar activity toward Erwinia carotovora and Ralstonia solanacearum, both potato pathogens (soft rot and bacterial wilt respectively).
Isolate A47 was identified as an endospore forming Gram positive bacterium associated to the genus Bacillus (Gordon et al. 1973; Norris et al. 1983). The bacteriological tests, thermal stability and chemical properties of the antibiotic products of this isolate suggests it as a B. subtilis strain (Silva,1992). Furthermore, a heat resistant compound obtained from a similar isolate has been partially purified and proven active against fungi. This compound was identified as a member of the iturin group (McKeen et al. 1986), and share the same characteristics of the antibiotic compound produced by isolate A47.
Iturin compounds constitute a family of lipopeptides excreted from several strains of B. subtilis when grown on liquid media. The common feature of these amphiphilic compounds is a peptide ring of seven aminoacid residues, including an invariable D-Tyr2, with the constant chiral sequence LDDLLDL, closed by a C14-C17 aliphatic b -aminoacid (Maget-Dana and Peypoux, 1994). They exhibit a strong antifungal activity against a wide variety of plant pathogenic fungi and yeast, but its antibacterial activity has not been deeply studied so far. Pusey and Wilson 1985 isolated a B. subtilis strain (B-3) from soil, active against the plant pathogen Monilinia fructicola. Later on, Besson et al. 1987 identified the presence of antifungal antibiotics of the iturin group in about 30 B. subtilis strains, and Besson and Michel 1987 identified two new types of iturins (D and E), differing from iturin A by the presence of a free carboxyl group in iturin D, and a carboxymethyl group in iturin E.
There is vast information about the selection of antagonistic microorganisms from nature, their effects, and the characterization of the metabolites involved. However, few data are available about the genetic alteration of the wild strains to improve their antagonistic effect. We believe that this is a powerful tool to produce new strains and products for biological control of plant pathogens, because the genetic modification using mutagenic agents is a simple technique and do not requires the insertion of foreign DNA in the antagonistic microorganism.
The objectives of this research were:
1) To select by random mutageneis of the wild type A47 Bacillus subtilis a strain with increased activity against the plant pathogens Botrytis cinerea, Ralstonia solanacerarum and Erwinia carotovora var. Carotovora.
2) To optimize culture conditions for the production of the antibiotic metabolite in submerged fermentation.
3) To isolate the metabolite with antibiotic activity. Materials and Methods Isolates. The bacterium, identified as Bacillus subtilis A47, was isolated by Silva (1992) and provided by the Plant Bacteriology Laboratory, Institute of Plant Production and Protection, Austral University of Chile, Valdivia, Chile.
The plant pathogenic microorganisms used as antibiotic targets were Botrytis cinerea VILL22 isolated from fresh raspberry fruits, (Silva, 1992), Erwinia carotovora var carotovora and Ralstonia solanacearum.
Induction, selection and characterization of bacterial mutants induced with acridine orange. To each one of eight tubes with 2 mL peptone, 0, 4, 8, 12, 16, 20, 24 and 40 µL of a 1mg/mL acridine orange solution were added. All tubes were inoculated with 50 m L from a primary culture of A47 strain, at a cell concentration equivalent to 1.0 O.D. at 600nm. The incubation was performed at 25ºC in an orbital shaker at 150 rpm for 24 h.
Samples from each tube were inoculated on CPG agar plates (10,0 gr/L glucose; 1,0 gr/L casaminoacids; 10,0 gr/L peptone; 15,0 gr/L agar-agar; pH 7.0) and used to obtain colonies and select potentially overproducing mutants. Plate tests were conducted by challenging selected colonies with cultures of B. cinerea, R. solanacearum and E. carotovora. Inhibition caused by isolate A47 was always used as control. The larger size of inhibition caused by mutant colonies was used as criterion to select overproducing colonies.
Some basic microbiological characteristics of selected mutant strains were conducted, such as Gram stain, glucose and lactose fermentation, growth on citrate, production of phenylalanine deaminase, indol production, and urea utilization. These tests were conduced in order to compare them with wild strain A47.
Selection of culture media and determination of bacterial growth of strain M40. Three different liquid culture media were used to study the level of production of inhibitory substances by overproducing isolate M40. The media were CPG (10 gr/L glucose; 1 gr/L casaminoacids; 10 gr/L peptone; pH 7.0), CPM (10 gr/L mannitol; 1 gr/L casaminoacids; 10 gr/L peptone; pH 7.0) and CPM-Ca2+ (10 gr/L mannitol; 1 gr/L casaminoacids; 10 gr/L peptone; 5 gr/L CaCl2 x 2H2O; pH 7.0). The best culture medium was selected according to the size of the growth inhibition zone of the pathogen in the agar plate.
The growth of strain M40 was carried out in CPM-Ca2+ broth at 200 rpm and 30ºC for 258 h. A primary culture of M40 with 2.5E+9 colony forming units (CFU) was inoculated in 500 mL of CPG-Ca2+ medium, and aliquots were taken between 6 and 12 h. Growth was determined by measuring the O.D. at 660 nm, sterile CPM-Ca2+ broth being used as blank.
When maximum bacterial growth was achieved, the culture was centrifuged at 10.000 x g during 30 min. The cellular pellet was discarded and the supernatant was acidified to pH 2.5 with 6M HCl.
The solution was autoclaved at 0,75 atm for 10 min and then centrifuged at 8.000 x g during 10 min, discarding the precipitate. The supernatant was adjusted to a final pH of 7.3 with phosphate buffer (1M, pH 8.0). This solution was designated as crude supernatant.
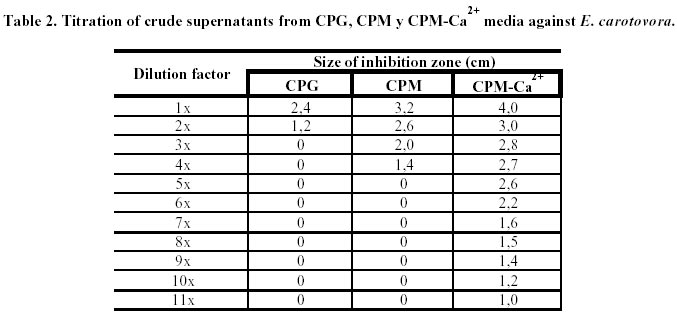
Determination of the inhibitory activity units (IAU) of the antibiotic metabolite in the crude supernatant. The crude supernatant obtained by liquid cultivation of isolate M40 was sequentially diluted with phosphate buffer (1M, pH 7.3) 2x, 3x, 4x, 5x, 6x, 7x, 8x, 9x, 10x, 11x and 12x times. Each of these dilutions was tested against E. carotovora in order to establish the minimum inhibitory concentration of the antibiotic metabolite against the plant pathogens. One IAU was defined as the higher dilution of the crude supernatant that inhibits in vitro the growth of the target phytopathogen.
Plate assay to establish the bacteriostatic or bactericidal activity of the antibiotic metabolite in the crude supernatant. From inhibitory assays conducted with E. carotovora and R. solanacearum, samples with sterile cotton tips were taken from the surface of the agar where no pathogen growth was observed. The tips were surface staked to CPG agar plates and incubated during 48 h. Growth or no growth, made the difference between bacteriostatic and bactericidal activity of crude supernatant from isolate M40.
Thermostability and solubility in methanol of the antibiotic metabolite in the crude supernatant. 1.5 mL of crude supernantants were dispensed in Eppendorf sterile tubes. The tubes were treated as follows: a) maintained at room temperature, b) water bath heated at 100ºC during 15 min and c) autoclaved at 120ºC during 20 min. From each set of tubes, fractions of 100 µl were tested against E. carotovora for inhibitory activity.
On the other hand, the antibiotic metabolite was extracted from the crude supernatant with one volume of methanol. Both fractions, soluble and insoluble, were evaporated to dryness, resuspended in distilled water and assayed against the phytopathogens.
Reverse liquid chromatography of the antibiotic metabolite. The methanol soluble fraction, obtained above, was chromatographed in a µ-Bondapack C-18 column (3,9 x 300 mm) balanced in 100% methanol. The samples were eluted with 100% methanol at a flow rate of 0,7 mL/min and a total time of 12 min. Peaks were collected, evaporated to dryness and assayed against R. solanacearum. The equipment was a SHIMADZU LC-10A coupled to a detector SPD-10AV. Procedures were as those previously reported (Schreiber et al. 1988; Gueldner et al. 1988).
Results Production of mutants using acridine orange as mutagen Twenty colonies isolated from strain A47 were mutated with acridine orange and more than twenty Petri dishes were produced containing numerous mutant colonies with different morphologies of growth on CPG agar. Colonies from each Petri dish were randomly selected with a sterile loop and tested for antagonistic activity. Five mucous colonies were selected with an increased activity with respect to the wild strain. Such strains were labeled as M40, M20, M16, M12 and M4. Results from inhibition tests against B. cinerea are presented in Figure 1a and Figure 1b, where the stronger antagonistic effect of all mutant strains with respect to wild strain is apparent. Strain M40 was selected for further studies because it produced bigger inhibition zones on B. cinerea (Figure 1b), Erwinia caratovora and Ralstonia solanacearum, and because it was easily cultivated and fast-growing on CPG, CPM and CPM-Ca2+ solid media.
Characterization of the selected mutant strain Mutant strain M40 was mucous-type and formed big, butyrous and round colonies on CPG agar. Under the microscope, cells were short bacilli forming irregular aggregates. As the wild strain, the mutant was Gram positive. Some biochemical tests revealed differences between the wild strain and mutant M40, the latter being able to grow on citrate as the sole carbon source (which reflects the presence of the citrate permease complex within the cell), and use phenylalanine as nitrogen source (which reflects the presence of phenylalanine deaminase). Results from such tests are in Table 1.
Selection of the proper culture media for the growth of the strain M40 Fermentation runs showed that mannitol was a better carbon source than glucose for antibiotic production, increasing the inhibition toward the pathogen twofold with respect to CPG medium. The addition of calcium ion to the CPM medium increased the antibiotic production in the mutant strain 5.5 and 2.5 fold with respect to the antibiotic production in the CPG and CPM media respectively.
From serial dilutions of the crude supernatants, volumetric activities against E. carotovora were 2, 4 and 11 IAU/100 m L for CPG, CPM and CPM-Ca2+ medium respectively. Adequate volume for inhibitory tests was 100 µL. Results are presented in Table 2.
Determination of the bacterial growth on the selected medium A primary culture of 2.5E+9 CFU was inoculated in 500 mL of CPG-Ca2+ medium. Sustained growth was observed for more than five days on CPG-Ca2+ medium, with a growth phase of more than eighty hours. Maximum cell growth was obtained at 114 hours, with 6.1E+8 CFU/mL. Synthesis of the antibiotic in the mutant strain started during the growth phase and maximum concentration was obtained after the end of that phase. Results are presented in Figure 2.
Thermostability and solubility in methanol of the antibiotic activity of the crude supernatant from mutant strain M40 Results on the thermal stability of the antibiotic are presented in Table 3. Different thermal treatments did not affect the antibiotic activity of the crude supernatant, inhibition zones being of the same size for aliquots subjected to the different treatments and also for untreated aliquots, used as control. Therefore, the antibiotic produced is thermoresistant, as it is the one produced by the wild strain A47. Similar results (data not shown) were obtained using the other phytopathogens as targets.
The antibiotic was soluble in methanol. The methanol soluble fraction was active against the phytopathogens, and no inhibition was obtained with the methanol insoluble fraction.
Reverse phase liquid chromatography of the antibiotic activity The HPLC analysis revealed three well-defined peaks (Figure 3). Samples from each peak were evaporated to dryness, resuspended with distilled water and assayed against R. solanacearum. Fraction number 2 in the chromatogram contained the antibiotic activity and was the major fraction with a retention time of 3,904 min. The other two peaks were devoid of antibiotic activity against the target phytopathogens. Discussion Previous studies demonstrated that a strain of Bacillus sp, labeled as A47, produced a thermostable metabolite active in vitro against the phytopathogens B. cinerea, E. carotovora and R. solanacearum. The metabolite was thermostable, protease resistant, soluble in methanol and classified as an antibiotic of the iturin group attending to its chemical and antibiotic properties (Silva, 1992).
In this work, wild strain A47 was subjected to mutation with acridine orange with the purpose of improving its antagonistic capacity towards phytopathogens. Acridine dyes are aromatic compounds that intercalate within pairs of bases in the DNA molecule favoring insertions and deletions of nucleotidic bases upon replication (Kapuscinski and Darzynkiewicz, 1984).
After mutation of wild strain A47 with acridine orange, five mutant strains were isolated with increased activity against the target phytopathogens. Among them, strain M40 was selected for further studies. This strain formed big, mucous colonies on CPG agar, quite different from the star-shaped colonies of the wild strain. Morphological differences were also revealed under the microscope, bacilli being much shorter in strain M40 than in the wild strain. Strains also performed differently in biochemical tests. Mutant strain M40 gave a positive phenylalanine test, revealing the presence of phenylalanine deaminase, which might reflect induction as a result of an alteration at the level of genes controlling transcription of the structural gene for the enzyme. Possibly, the structural genes are negatively regulated in the wild strain, blocking enzyme synthesis. Mutant strain M40 was able to grow on citrate as the sole carbon source, revealing the presence of the permease complex for citrate transport into the cell (Davis et al. 1980), while the wild strain gave a negative test for citrate utilization. Despite this, the inhibition spectrum for target phytopathogens of the wild strain did not change upon mutation.
Fermentation kinetics of the mutant strain M40 exhibited a prolonged growth phase, reaching an equivalent of 6.1E+8 CFU/mL at 114 h of cultivation. The synthesis of the antibiotic begins during growth phase, as proved by inhibition tests performed with crude supernatants from mutant strain M40 harvested from 80 to 120 hours, but maximum concentration was obtained after the growth phase. Synthesis of peptide antibiotics usually starts at the end of exponential growth, reaching maximum concentration after cell growth has ceased (Bodanzky and Perlman, 1969). It has been suggested that many microorganisms can synthesize antibiotics while in growth phase (Haavik and Thomassen, 1973; Barr, 1975; Haavik, 1976), which is in agreement with production kinetics in mutant strain M40. The factor triggering the onset of antibiotic synthesis is more likely the exhaustion of a limiting nutrient required for cell growth. This limitation usually stimulates differentiation which, for the case of bacilli, means endospore formation. Sporulation is associated with the synthesis of a new cell wall and the degradation of that in the mother cell. This cell wall, as the antibiotic, contains D-aminoacids, and it has been suggested that an altered metabolism for cell wall synthesis could furnish the precursors for antibiotic synthesis
We found that the synthesis of the antibiotic was increased when the mutant strain M40 was cultured on CPM medium, which contains mannitol instead of glucose as carbon source. This is in agreement with Besson et al. 1987, who reported mannitol, fructose and sucrose as better carbon sources than glucose for the synthesis of iturin A in B. subtilis. Catabolite repression is a regulatory mechanism by which the cell coordinates the metabolism of the carbon and energy source, to maximize the efficiency of nutrient utilization and control other metabolic processes. It is a well-documented fact that glucose promotes repression of the enzymes involved in the catabolism of the carbohydrates metabolized at lower rates. As a result, a hierarchy of utilization of carbon and energy sources is established. According to this, the synthesis of secondary metabolites is subjected to catabolite repression by glucose, either directly or indirectly (Fischer and Sonenshein, 1991).
Calcium ion at a level of 0.5% had a significant effect on antibiotic production by the mutant strain M40 on CPM medium. A similar effect was observed in a culture of Lactococcus lactis IO-1, where the addition of calcium ion to the medium produced a significant increase in the production of the antibiotic nisin Z (Matsusaki et al. 1996). However, an opposite effect was observed by Besson et al. 1987, who reported that the addition of CaCl2 to the culture medium of a B. subtilis strain, inhibit the excretion of iturin A completely. They postulate that Ca++ modifies the permeability of the cell membrane in Bacillus blocking the excretion of iturin A. However, later on, the same authors proved that Ca2+ and Mn2+ promoted iturin A and bacillomycin L precipitation after excretion into the medium (Besson and Michel, 1991). In our case, the increase in concentration of antibiotic in the medium can be a consequence of the increase in cell wall permeability of mutant strain M40 promoted by Ca2+, which is in agreement with Petit-Glatron et al. 1993, who studied the capacity of the cell wall to concentrate Ca2+ and proposed that the increased concentration in the microenvironment of the cell wall could play an important role in the last step of the secretion.
A bioassay based on clearing zone on agar plate was used to determine the antibiotic activity of the metabolite produced by the mutant strain M40 against the phytopathogens B. cinerea, E. carotovora and R. Solanacearum. Inhibition zones revealed the production of a strong antagonist by the mutant strain M40. These results are interesting, because the antibiotic produced inhibits growth of both prokaryotes and eukaryotes, as is the case for other iturins synthesized by B. subtilis (Maget-Dana and Peypoux, 1994). According to our results, the antibiotic produced by mutant strain M40 has bactericidal and probably fungicidal activity. The target site for iturin A and bacillomycin L on yeast cells is in the plasma membrane. Both antibiotics promote spheroplast lysis and rapidly increase permeability to K+, which has been associated with its fungicidal activity (Besson et al. 1984). The ability of iturin antibiotics to increase membrane permeability of target microorganisms is due to the formation of ion channels on the cell membranes. In this context, a tertiary structure iturin-phospholipid-sterol was suggested to be the biologically efficient structure (Maget-Dana and Peypoux, 1994).
The different thermal treatments conducted with supernatants from cultures of the mutant strain M40, demonstrate the thermoresistance of the inhibitory metabolite. Growth inhibition zones produced in B. cinerea, E. carotovora and R. solanacearum plates were the same for all supernatants, independent of the thermal treatment to which they were subjected. These results are in agreement with the reported thermoresistance of iturin antibiotics produced by Bacillus.
With respect to the solubility of supernatants, they differ though not significantly of those obtained for antibiotics isolated from different Bacillus species. The metabolite produced by mutant strain M40 was soluble in water and methanol. Iturin A and bacillomycin L were soluble in methanol and water-methanol mixtures too (Besson et al. 1977; Peypoux et al. 1978). On the other hand, bacillomycin D is insoluble in water and in most organic solvents (Peypoux et al. 1980). A new antifungal peptidolipid antibiotic, iturin AL, isolated from a strain of B. subtilis, was soluble in methanol and ethanol and barely soluble in water, enough though to allow its detection by diffusion in agar (Winkelmann, 1983). Later, Schreiber et al. (1988) isolated from Ulmus americana a metabolite of 500 D, soluble in water, methanol, ethanol and chloroform, quite similar to the antifungal metabolites described by McKeen et al. (1986).
The strategy for isolation and purification of the antibiotic produced by the mutant strain M40 was reverse phase HPLC. A simple chromatogram was necessary by this procedure; only three peaks were obtained and the antibiotic activity was in only one of them. This procedure has been previously used to purify other antibiotics (Azevedo et al. 1993; Razafindralambo et al. 1993; Eshita et al. 1995; Kajimura et al. 1995).
Further research is needed to determine the chemical structure of the antibiotic produced by the mutant strain M40 within the family of iturins. Another aspects, which require further work, is a thorough study on the kinetics of antibiotic production and the optimization of medium design and product recovery. More knowledge is required on the mechanisms of biocontrol of phytopathogens to develop rational strategies for the application of the antagonists and their metabolites within the agroecosystem. Once elucidated such strategies, genetic engineering can provide an efficient way of gathering desirable characteristics from different organisms in only one organism ecologically adapted to a particular system. This can be accomplished by cloning the biosynthetic genes in a competent organism like Rhizobium or other members of the rhizosphere. The cloning of such genes directly into the plant cells is also interesting option. There is still a long way to go before a sound systems is developed to protect plants from their predators without altering the ecological balance among species. The use of bioantagonists is certainly a very promising route. References
Note: EJB Electronic Journal of Biotechnology is not responsible if on-line references cited on manuscripts are not available any more after the date of publication. Supported by UNESCO / MIRCEN network © 2002 by Universidad Católica de Valparaíso -- Chile The following images related to this document are available:Photo images[ej02009t3.jpg] [ej02009f1.jpg] [ej02009f3.jpg] [ej02009t2.jpg] [ej02009f2.jpg] [ej02009t1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}