 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Electronic Journal of Biotechnology, Vol. 5, No. 1, April, 2002 Efficient transformation of Penicillium chrysogenum mediated by Agrobacterium tumefaciens LBA4404 for cloning of Vitreoscilla hemoglobin gene Chuan-Bao Sun*#1, Qiu-Lian Kong2 and Wen-Si Xu3 1
of Genetic Engineering, Department of Bio-pharmaceutical, Shanghai Institute of
Pharmaceutical Industry, Shanghai, 200040 P. R. China E-mail: chuanbaosun@hotmail.com Financial support: Shangai Postdoctoral Financial Comitee. Received
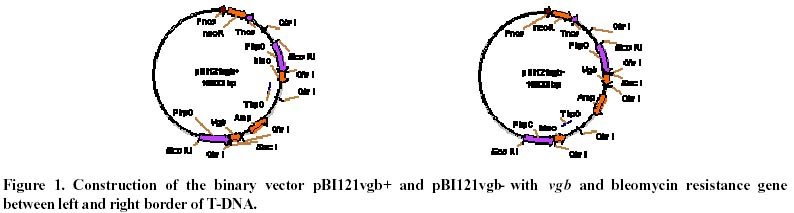
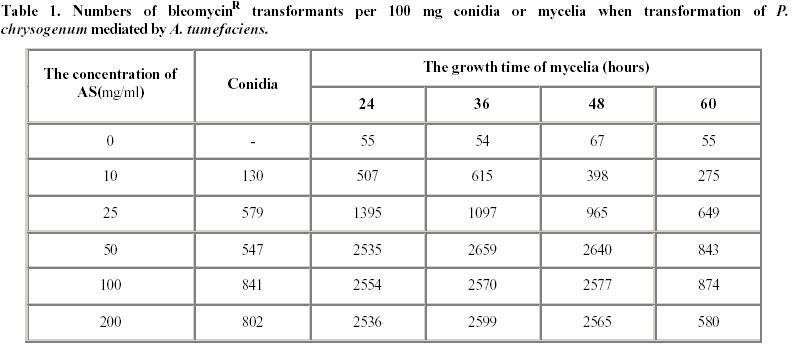
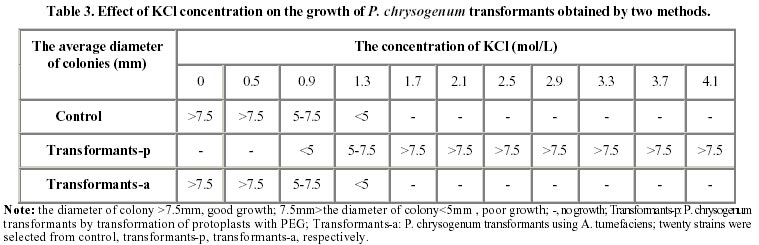
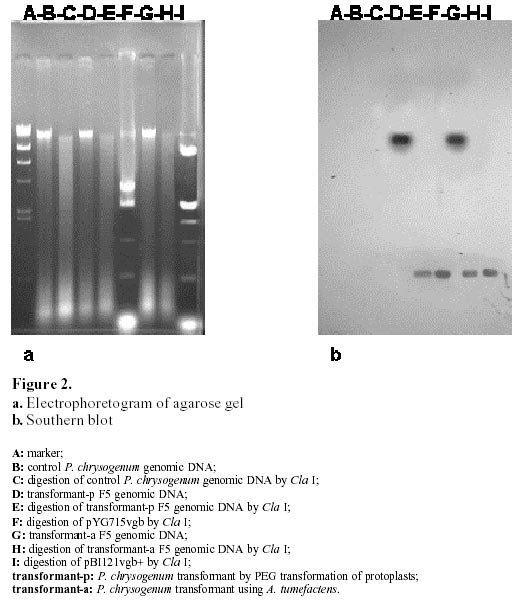
October 8, 2001 Code Number: ej02010 Abstract The vgb and bleomycin resistance genes could be efficiently transferred into the filamentous fungus Penicillium chrysogenum under help of T-DNA of Agrobacterium tumefaciens LBA4404 and the transferred genes were integrated at a chromosomal locus of P. chrysogenum. Transformation using A. tumefaciens LBA4404 could be enhanced in the presence of acetosyringone (AS) when being carried out in the conidiaand mycelium. The efficiencies of transformation could be improved up to 10 folds compared with conventional techniques for transformation of P. chrysogenum protoplasts. Keywords: Agrobacterium tumefaciens, Penicillium chrysogenum, Ti plasmid, transformation. Article Penicillins are b-lactam antibiotics which have broad clinical utility. Penicillins are produced by a number of microorganisms, including the several fungal species and several streptomycete species. Penicillium chrysogenum is an important industrial organism due to its capacity to produce penicillin which is still one of the main commercial antibiotics (Brakhage, 1998). For decades, penicillin yields have been increased through development of better production strains by classical mutagenesis procedures and optimization of the growth conditions (Vanek and Hostalek, 1986; Fernandez-Canon et al. 1989; Keshavarz et al. 1989). Currently, together with molecular techniques, this facilitated a thorough analysis of the genetic regulation of metabolic pathways. The great progress in elucidation of the molecular regulation of biosyntheses of penicillin has been made in the penicillin producer P. chrysogenum. Since the biochemistry of penicillin biosynthesis is rather well understood and recombinant techniques have been developed for some filamentous fungi, the overexpression of biosynthesis genes will lead to higher yields of penicillin in the respective production strains. Therefore, recombinant DNA technology of filamentous fungus will be an important tool for future strain improvement strategies with P. chrysogenum (Carr et al. 1986; Smith et al. 1990; Fernandez-Canon and Penalva, 1995). Penicillin biosynthesis is regulated by environmental factors such as the phosphate, carbon, nitrogen and oxygen content of the medium (Feng et al. 1994). The overall rate of penicillin synthesis is severely reduced under conditions of low oxygen. Reduction of oxygen supply leads to accumulation of penicillin N, a precursor of penicillin. The mechanism of oxygen control over penicillin synthesis is not well understood. Possibly, low oxygen levels directly affect the biosynthetic pathway of penicillin, which includes several oxidation reactions. It is also possible that a more efficient overall metabolism provided by higher oxygen levels indirectly results in higher penicillin yields. Regardless of the mechanism, technologies that improve aerobic metabolism in these organisms should have a positive effect on penicillin production. Vitreoscilla hemoglobin (VHb) is so far the only one found in prokaryote. The expression of VHb is controlled by oxygen, and the amount of VHb expression increases greatly in lower oxygen environment. Possible mechanisms for VHb action include increasing the flux of oxygen to the respiratory apparatus, providing higher internal oxygen concentrations, altering the internal redox state, or functioning as an efficient terminal oxidase. VHb plays a major role in improving the conditions of bacterial growth, increasing the synthesis of protein and secondary metabolites, especially antibiotics, as well as promoting expression of heterogenous genes. The characteristic of VHb binding oxygen provides for us a possibility to enhance both the oxygen transfer and the efficiency of oxygen absorption in cells. Intracellular expression of a bacterial heme-binding protein (Vitreoscilla hemoglobin, or VHb) has resulted in higher productivity of a number of industrial cell types. For example, the expression of VHb in the filamentous bacterium Streptomyces coelicolor resulted in tenfold higher yields of the polyketide actinorhodin in bench scale batch fermentations run under reduced aeration (Magnolo et al. 1991). Also, the efficiency of cloned protein synthesis by oxygen-limited Escherichia coli cultures was increased in VHb-expressing strains (Khosla et al. 1990). Several VHb-expressing transformants of Acremorium chrysogenum produced significantly higher yields of cephalosporion C than control strains in batch culture experiments (Demodena et al. 1993). Therefore, intracellular expression of VHb in P. chrysogenum by recombinant DNA technology could provide a possibility that a productivity of penicillin was increased. Thus a basic requirement to establish a recombinant DNA system for the cloning of vgb gene (coding VHb) in P. chrysogenum is a gene transfer system. This includes a reproducible transformation procedure and proper selection markers. Several transformation techniques are available for filamentous fungi. Most protocols involve the transformation of protoplasts using electroporation or by a combination of CaCl2 and polyethylene glycol (PEG). Filamentous fungi were transformed using two dominant selection markers, namely the bacterial resistance gene and complement gene of auxotrophic (Sanchez et al. 1987; Kolar et al. 1988). Intact fungal cells have also been transformed by the transformation of protoplasts, but usually with a lower transformation frequency. Agrobacterium tumefaciens can transfer part of its Ti plasmid, the T-DNA, into plant cells during tumorigenesis (Winans, 1992; Hooykaas and Beijersbergen, 1994; Ishida et al. 1996). It is routinely used for the genetic modification of a wide range of plant species. For years, the transformation using A. tumefaciens has been applied to the yeast Saccharomyces cerevisiae and filamentous fungi, including Aspergillus niger, Fusarium venenatum, Trichoderma reesei, Colletotrichum gloeosporioides, Neurospora crassa, and the mushroom Agaricus bisporus (Bundock et al. 1995; Bundock and Hooykaas, 1996; de Groot et al. 1998). Here we report that Agrobacterium tumefaciens LBA4404 transfers part of its Ti plasmid pBI121vgb+, the T-DNA with vgb and bleomycin resistance gene, to P. chrysogenum cells. The T-DNA then integrates into the P. chrysogenum nuclear genome at a random position. Materials and Methods Strains. Fungal strain P. chrysogenum was obtained from biology department of Shanghai institute of pharmaceutical industry. Escherichia coli strain DH5 a was used as recipient hosts for all DNA work. The A. tumefaciens strain LBA4404 was used for the transformation of the P. chrysogenum. Plasmid construction. pBI121vgb+ was constructed as follows. A 3kb SacI/HindⅢ fragment containing Gus gene was deleted from the binary vector pBI121, the remaining other 10kb large fragment containing T-DNA sequence was blunted and ringwise ligated itself. The EcoR I site was between left and right border of T-DNA. Meantime a 6.3 kb EcoR I frangment was obtained from pYG715/vgb, which containing the vgb structural gene and bleomycin resistance gene, both were fused to the transcriptional promoter region of the trpC gene of A. nidulans (PtrpC). Then the 6.3kb EcoR I fragment was ligated into the EcoR I sites between left and right border of T-DNA of 10kb large fragment from the binary vector pBI121, resulting in pBI121vgb+ and pBI121vgb-. Media and conditions. For the penicillin production experiments, seed cultures were prepared as follows. Spores from P. chrysogenum clones were grown on LCS Agar {lactose, 1.5%(w/v); corn-steep liquor, 0.25%(v/v); peptone, 0.5%(w/v); sodium chloride, 0.4%(w/v); magnesium sulfate, 0.05%(w/v); potassium dihydrogen phosphate, 0.06%(w/v); ferric chloride, 0.0005%(w/v); cupric sulfate, 0.0002%(w/v); agar 3.0%(w/v)}, harvested, and inoculated into 25 ml defined seed media {glucose, 3.0%(w/v); pharmamedia, 1.0%(w/v); corn steep liquor, 3.0%(v/v); ammonium sulfate, 0.2%(w/v); calcium carbonate, 0.5%(w/v); potassium dihydrogen phosphate, 0.05%(w/v); lactose, 1.0%(w/v); primary dry yeast, 1.0%(w/v)} in a 250 ml glass flask. The seed cultures were incubated at 25ºC and 220 rpm for 48 hours in an orbital incubator. 5 ml of the seed culture was inoculated into 25 ml of minimal production media {lactose, 12.0%(w/v); pharmamedia, 2.5%(w/v); ammonium sulfate, 1.05%(w/v); calcium carbonate, 1.0%(w/v); calcium sulfate, 0.5%(w/v); sodium sulfate, 0.4%(w/v); lard oil, 1.0%(w/v); potassium dihydrogen, 0.05%(w/v); potassium sulfate, 0.5%(w/v); phenoxyacetic acid, 1.25%(w/v)} in 250 ml glass flasks. The cultures were incubated at 28ºC and 220rpm for 240 hours (Vanek and Hostalek, 1986b). P. chrysogenum transformation. P. chrysogenum conidia were obtained by growing the strain on LCS Agar plate for 5 days and washing the conidia with a physiologic salt solution. Cocultivations between A. tumefaciens and P. Chrysogenum was performed as follows. For transformation of conidia, 100 ml conidia at a concentration of 106-108 conidia/ml were mixed with 100 ml of the A. tumefaciens culture prepared as described. The A. tumefaciens containing the binary vector pBI121vgb+ were obtained by growing the strain on a LB plate for 2-3 days and washing the strain with a physiologic salt solution. When mycelia were transformed, 100 ml mycelia at a concentration of 1 gram/ml were used. Subsequently, these mixtures incubated into 50 ml defied seed media with or without acetosyringone (AS) at 25ºC and 220rpm for 48 hours. Then these mixtures cultures were transferred to P. chrysogenum minimal medium plates containing 10 mg/ml cefotaxime to kill the A. tumefaciens cells and 20 mg/ml bleomycin to select for transformants. Transformation of P. chrysogenum was performed using the polyethylene glycol method as described previously (Cantoral et al. 1987). Isolation of DNA and Southern hybridization. The vegetative mycelial growth from a 48 hour culture prepared as descrebed above was collected by filtration through cheesecloth, frozen in liquid nitrogen and lyophilized overnight. The dried mycelia were groud with sand in a mortar and pestle and resuspended in 25 ml of 100 mmol/L LiCl, 50 mmol/L EDTA, 10 mmol/L Tris pH8.0, 4% SDS. After heating the suspension to 55ºC in a 60ºC water bath, the mixture was extracted first with 1 mol/L Tris (pH 8.0) saturated phenol, followed by Tris-saturated pheonl:chloroform (1:1, v:v) and then chloroform RNA was precipitated from the aqueous phase by the addition of an equal volume of cold 6mol/L LiCl and then the mixture was remained at –20ºC for two to three hours. After centrifugation at 12000 g for 20 minutes at 4ºC, the supernatant was made 66% (v/v) ethanol and cooled to –20ºC for 15 minutes to precipitate the DNA. After centrifugation as described above, the DNA pellet was washed with 70% ethanol, dried and resuspended in TE Buffer (10 mmol/L Tris-HCl, pH 7.5, 1 mmol/L EDTA) (Conder, 2000). The DNA concentration was estimated by comparison to known DNA standards when stained with ethidium bromide in agarose gel electrophoresis. 5 mg of the DNA samples digested by Cla I were run on a 0.7% agarose denaturing gel and blotted to nitrocellulose filter. The filter was hybridized to probes with the vgb gene probe. The blotting and hybridizations were carried out under standard conditions. Results Construction of the binary vector The binary vector was constructed for transfer T-DNA from A. tumefaciens to P. chrysogenum. The vectors, pBI121vgb+ or pBI121vgb- (Figure 1), were constructed based on the bidirectional integration of the vector pYG715vgb and the vector pBI121. The vgb gene and the bleomycin resistance gene fused downstream of the transcriptional promoter region of the trpC gene from Aspergillus nidulans (PtrpC) and were between left and right border of T-DNA of the binary vector. Effect of the acetosyringone concentration and the mycelia collected at different growth time on transformantion of P. chrysogenum mediated by A. tumefaciens In order to establish a system in which A. tumefaciens could transfer T-DNA to the filamentous fungus P. chrysogenum, we constructed the binary vector pBI121vgb+. This plasmid contains the vgb and the bleomycin resistance gene between the left border and right border repeats of the binary vector pBI121. Plasmid pBI121vgb+ was then electroporated into the A. tumefaciens LBA4404 helper strain, which carries the vir genes that code for the T-DNA transfer system. Incubation of P. chrysogenum cells with A. tumefaciens could lead to the formation of bleomycin-resistant colonies, and a greater number of bleomycin-resistant colonies were found if the cocultivation was done on medium containing AS (Table 1). The negative controls never gave rise to resistant colonies. This result shows that induction of the vir genes is essential for transfer of the resistance trait to the mold cells and wherefore suggests that A. tumefaciens can use its virulence system to transform the filamentous fungus P. chrysogenum. The transformation frequency varied from approximately 50 to 2700 transformants per 100 mg conidia or mycelia. Comparison of transformation using A. tumefaciens with the transformation of protoplasts The transformation frequency using A. tumefaciens is up to 10 times higher than the values obtained for PEG transformation of protoplasts (Table 2). These transformants obtained by both transformation methods showed different growth in the medium containing different KCl concentration (Table 3). Transformants-p get a halophilism, and transformants-a show same growth with control strain in the medium containing different salt concentration. We presumed that the DNA transfer mechanism is different in the two transformation methods. The stability of bleomycin resistance in P. chrysogenum transformants The stability of the bleomycin resistant phenotype was confirmed. First, transformants obtained by transformation of protoplasts and by A. tumefaciens were transferred to fresh plates without bleomycin and the transfer were carried out from one generation to the next until the fifth generations. Then transformants F1-F5 were transferred to fresh plates containing 20 mg/ml bleomycin. The results showed that the bleomycin resistant phenotype of transformants obtained by both transformation methods was heritable and stable (Table 4). Then genomic DNA of the transformants F5 resistant to bleomycin were isolated and tested by Southern blotting analysis with vgb probe for vgb cloning (Figure 2). Therefore the vgb and bleomycin resistance genes were integrated into P. chrysogenum chromosome genome. Discussion The results demonstrated that the Agrobacterium tumefaciens LBA4404 could transfer its T-DNA into Penicillium chrysogenum. The transfer could depend on the induction of the bacterial virulence genes, which expression could mediate transfer of T-DNA into P. chrysogenum. A. tumefaciens can transform not only the conidia of P. chrysogenum, but also hyphal tissue. Transformantion using A. tumefaciens LBA4404 could be enhanced in the presence of acetosyringone when being carried out in the conidia and mycelia. However, acetosyringone is not necessary for transformation of DNA in mycelia. This paper showed that the efficiencies of transformation using A. tumefaciens could be improved up to 10 folds compared with the transformation of P. chrysogenum protoplasts combined of CaCl2 and polyethylene glycol. Transformants of P. chrysogenum obtained by transformation of protoplast got a halophilism, they could grow at presence of high concentration of KCl salt, but transformants obtained by A. tumefaciens couldn't. We presumed that the DNA transfer mechanism was different between two transformation methods. Because of A. tumefaciens LBA4404 can transfer a foreign DNA into conidia and mycelia of P. chrysogenum directly, this is a powerful and simple tool for genetic transformation of industrial P. chrysogenum. Finally, the successful cloning of vgb in P. chrysogenum by A. tumefaciens T-DNA transfer permits this technology to be applied in improving of industrial penicillin-producing strains as well. References
Note: EJB Electronic Journal of Biotechnology is not responsible if on-line references cited on manuscripts are not available any more after the date of publication. Supported by UNESCO / MIRCEN network © 2002 by Universidad Católica de Valparaíso -- Chile The following images related to this document are available:Photo images[ej02010f2.jpg] [ej02010t3.jpg] [ej02010t1.jpg] [ej02010t4.jpg] [ej02010t2.jpg] [ej02010f1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}