 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
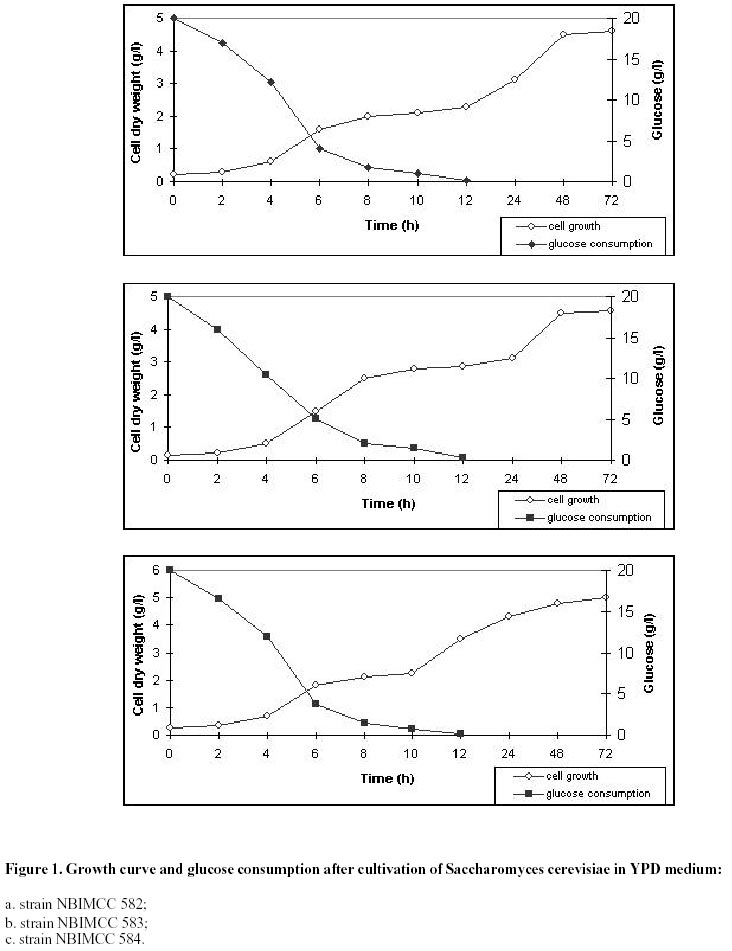
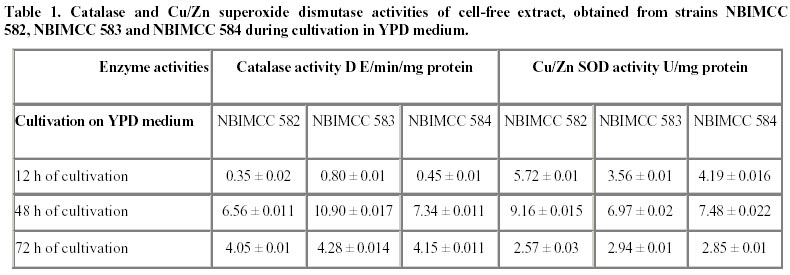
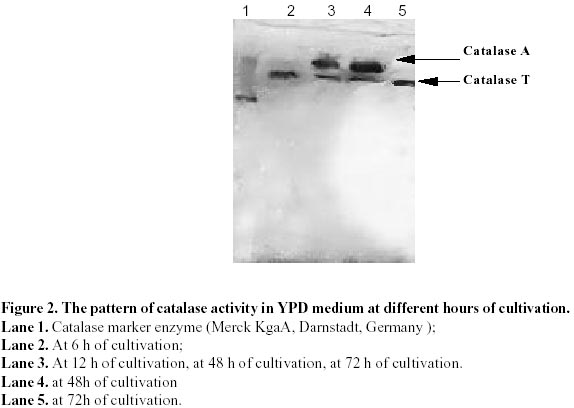
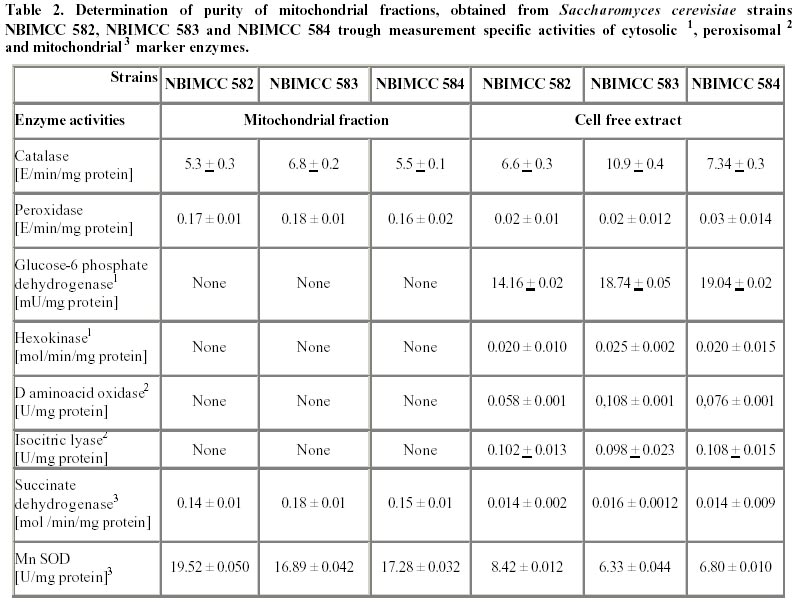
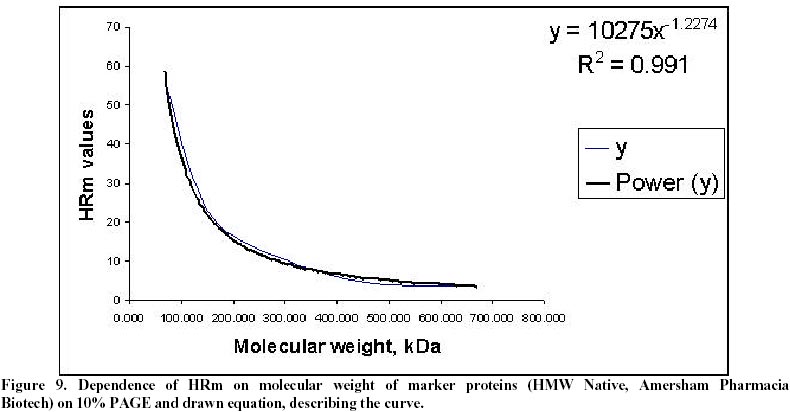
Electronic Journal of Biotechnology, Vol. 5, No. 1, April, 2002 Catalase enzyme in mitochondria of Saccharomyces cerevisiae Ventsislava Yankova Petrova1, Tanya Vassileva Rasheva2 and Anna V. Kujumdzieva* 3
1Department
of Cytology, Histology and Embryology, Biological Faculty, Sofia University "St.
Kliment Ohridski", 8 Dragan Tzankov Blvd., Sofia 1421, Bulgaria Tel: 359
2 63 30 277 E-mail: petrova_v@hotmail.com
Received December
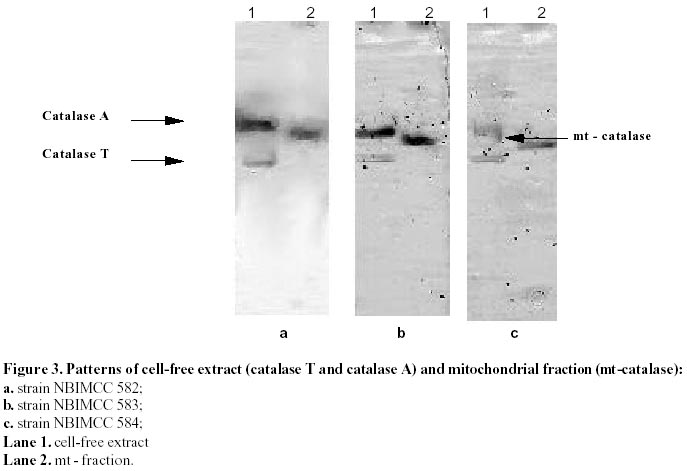
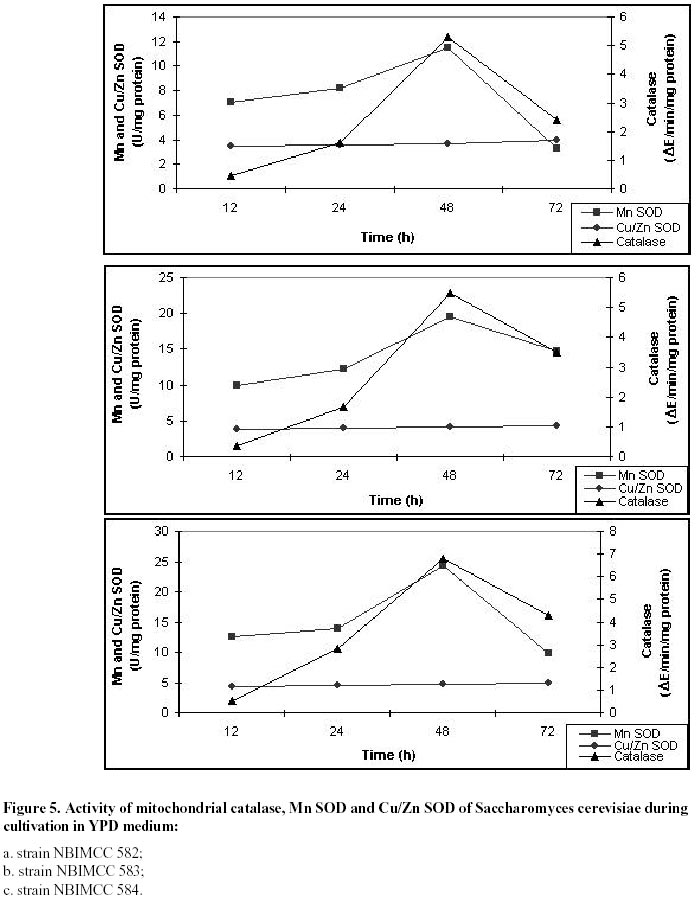
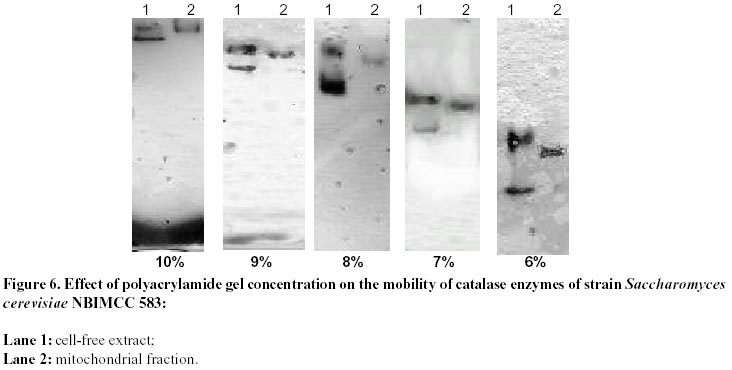
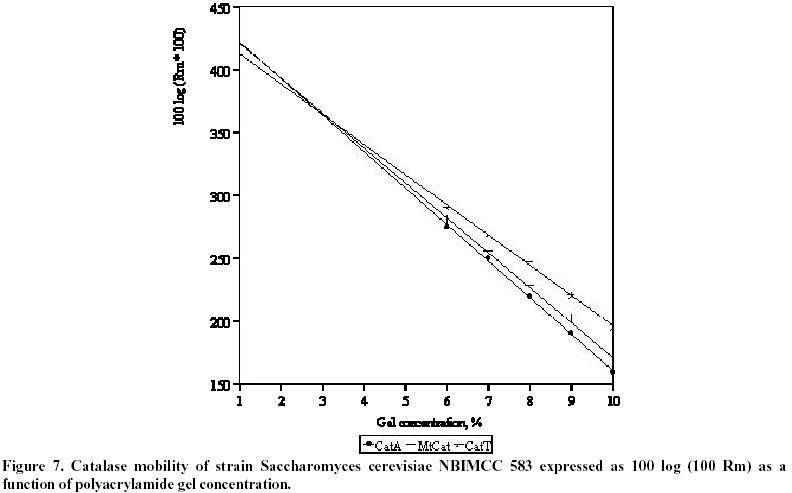
26, 2001 Code Number: ej02011 Abstract Catalase and superoxide dismutase activities have been explored in the yeast Saccharomyces cerevisiae during batchwise growth experiment. During the diauxic growth in YPD medium high Ys values were obtained (0.415 - 0.423) and correlation between the total activities of both enzymes has been found. A mitochondrial fraction from three type strains of Saccharomyces cerevisiae has been isolated. The purity of this fraction was proved through different enzyme assays: hexokinase, glucose-6-phosphate dehydrogenase, D-amino acid oxidase, isocitric lyase, succinate dehydrogenase. Then the catalase, peroxidase, Mn and Cu/Zn superoxide dismutase activities were evaluated in the mitochondrial fraction. Polyacrylamide gel electrophoresis separations allowed to identify a mitochondrial catalase as a band of 0.239 Rm value. It differed from the two catalase specific bands with Rm values 0.218 and 0.257 obtained from the crude extract. It was proved that the three catalase proteins are charge isomers. A positive correlation between the activity of mitochondrial catalase and Mn superoxide dismutase also takes place. Molecular weight of mitochonrial catalase protein has been determined as 240 kD. Keywords: catalase, mitochondria, Saccharomyces cerevisiae, SOD. Abbreviations: SOD: superoxide dismutase; DAB - 3,3’: diaminobenzidine tetrachloride; mt: mitochondrial; MW: molecular weight. Article Catalase is an essential enzyme in the decomposition of intracellular hydrogen peroxide (H2O2). In Saccharomyces cerevisiae cells, two different types of catalase have been found (Traczyk et al. 1985; Skoneczny and Rytka, 2000), which were designated catalase type A and catalase type T (Seah et al. 1973). The two enzymes are encoded by different genes (CTA1 and CTT1 respectively), possessing independent control and being localized in different compartments: catalase T is a cytoplasm enzyme whereas catalase A is localized in peroxisomes (Lapinskas et al. 1993). The molecular weights of the biologically active, homotetrameric enzymes are 170-190 kD and 225-250 kD for catalase A and T respectively (Klei et al. 1990). Studies with wild type Saccharomyces cerevisiae in continuous cultures on mixture of glucose and H2O2 indicated an enhanced level of mitochondrial cytochrome c peroxidase (Verduyn et al. 1988). These results suggested that mitochondria possess an independent mechanism for decomposition of exogenous H2O2 (Jamieson, 1995). They are also site of endogenous O2- production via autooxidation of ubisemiquinone and electron transport proteins and H2O2 formed by its spontaneous or enzymatic dismutation (Estruch, 2000; Kowaltowski et al. 2001). The scavenge of these reactive oxygen species is realized by mitochondrial antioxidant system, represented by Mn containing superoxide dismutase, Cu/Zn one found in the intermembrane space of the mitochondria (Ravindranath and Fridovich, 1975; Sturtz et al. 2001) and cytochrom c peroxidase (Charizanis et al. 1999). This well known mechanism for detoxification was added by Scandalios et al. 1980 who proved the presence of catalase enzyme in mitochondria of mesophyll cells of Nicotiana sylvestris and Radi et al. 1991 - in rat heart mitochondria. This evidence motivated investigations for presence of catalase in yeast mitochondria, besides of well known cytochrom c peroxidase degrading hydrogen peroxide. It could be expected that such kind of co-operative action of the above-mentioned H2O2 degrading enzymes is also possible. These observations directed us to investigate mitochondria for the presence of catalase activity in three different strains of Saccharomyces cerevisiae. Our results clearly indicated that a mitochondrial catalase enzyme is present in vivo and suggested the existence of an additional cellular compartment for localization of this protein in yeast cell. Materials and Methods Strains and growth conditions. The following strains of Saccharomyces cerevisiae obtained from the National Bank for Industrial Microorganisms and Cell Cultures were used: NBIMCC 582, NBIMCC 583 and NBIMCC 584. Cells were grown on a rotor shaker (204 rpm) at 30oC on standard liquid YPD medium (2% Glucose, 1% Yeast extract and 1% Bacto-Peptone). Preparation of cell-free extracts. Cells from 6, 12, 48 and 72 h of cultivation were harvested by centrifugation at 800 x g for 10 minutes and washed twice with distilled H2O. Cell wall disruption was carried out by zymolyase according to the procedure of Defontaine et al. 1991. The cell debris was removed by centrifugation at 1000 x g for 10 min. Then the cell free extracts were frozen and thawed 3 times in order to disrupt mitochondria and centrifuged at 15 000 x g for 10 min. Obtained supernatants were used for enzymatic analyses. Isolation of mitochondrial fraction. The fraction of heavy mitochondria was isolated from obtained spheroplasts after osmotic shock in distilled water applying the procedure of Holtta, 1977. The cell debris was harvested at 1000 x g for 10 min and the heavy mitochondrial fraction was isolated by centrifugation at 3500 x g for 20 min. Then the crude mitochondrial pellet was carefully washed with buffer, containing 0.5 M sorbitol, 50 mM Tris, 10 mM EDTA, pH 7.5. This procedure was repeated three times and collected mitochondria were resuspended in desionized water and lysed by freezing and thawing. The obtained sample of disrupted mitochondria was subjected to further biochemical analyses. Enzyme assays. All enzyme assays were performed in cell-free extracts as well as in mitochondrial fractions. Catalase [EC: 1.11.1.6] activity was determined spectrophotometrically according to Aebi, 1970. One unit of enzyme activity was expressed as D E/min/mg protein. Peroxidase [EC: 1.11.1.7] activity was assayed by the 3,3’ – diaminobenzidine tetrachloride [DAB] method of Selander et al. 1986. One unit of enzyme activity was expressed as D E/min/mg protein. Superoxide-dismutase [EC: 1.15.1.1] activity was measured as described by Beauchamp and Fridovich, 1971. One unit was defined as the amount of enzyme causing 50% decrease in the reduction of Nitro Blue Tetrazolium (NBT). The Mn – containing type of the enzyme was proved by inhibition analysis with 2 mM KCN (Geller and Winge, 1984). The succinate dehydrogenase [1.3.99.1] activity in mitochondrial fraction was determined by the method of Bonner, 1955. One unit of enzyme activity was expressed as mol reduced K3Fe(CN)6/min/mg protein. Hexokinase [2.7.1.1] activity in cell-free extract and mitochondrial fraction was defined according to Greene, 1969. One unit of enzyme activity was expressed as mol/min/mg protein. Glucose-6-phosphate dehydrogenase [1.1.1.49] activity was determined by the method of Greene, 1969. One unit of enzyme activity was expressed as mmol reduced NADP/min/mg protein. D-amino acid oxidase [1.4.3.3] activity was measured as described by Lichtenberg and Wellner, 1968. One-tenth unit of activity yields an absorbency of 0.180 at l =300 nm. Isocitric lyase [4.1.3.1] activity was measured as described by Daron and Gunsalus, 1962. One unit is defined as that amount of enzyme catalyzing the formation of 1 mmol of glyoxylate in 5 minutes at 28ºC, at pH 6.0. Analysis of glucose. Glucose was determined by the method of Somogy, 1952 and Nelson, 1944. Cell dry weight estimation. Cell dry weight was determined gravimetrically after drying washed cells to constant weight at 105ºC. Determination of kinetic parameters. Growth yield (Ys) was calculated as described by Pirt, 1975. Protein determination. Protein content was determined by the method of Lowry et al. 1951. Bovine serum albumin (Sigma-Aldrich Chemie GmbH, Germany) was used as a standard. Polyacrylamide gel electrophoresis. PAGE was performed on 10% resolving and 3.5% stacking gels according to Davis, 1964 on vertical electrophoretic unit (Mighty Small II Hoeferâ, Amersham Pharmacia Biotech). Catalase activity was visualized by incubating the gels in solution, containing 0.15M NaSO3 and 0.3M H2O2 for 15 min at room temperature, followed by brief rinsing with distilled water and further treatment with iodine solution (% KI and % I2). As a result dark blue bands of catalase activity on pale-blue background appeared. Peroxidase activity on acrylamide gel was developed using the method of Selander et al. 1986. Determination of charge isomers was performed following the methodology of Hedrick and Smith, 1968. Molecular weight determination of mitochondrial catalase was performed on 10% PAGE. As standard, proteins with known molecular weight - thyroglobulin 669 000, ferritin 440 000, catalase 232 000, lactate dehydrogenase 140 000, bovine serum albumin 67 000 (HMW Native, Amersham Pharmacia Biotech), stained with Coumassi blue were used. The HRm values of molecular markers were plotted against their molecular weight and equation, describing the curve was drawn. The molecular weight of the mitochondrial catalase was calculated, using the obtained equation. Results and Discussion Catalase activity and electrophoretic profile of the enzyme The catalase enzyme has been investigated during cultivation of Saccharomyces cerevisiae NBIMCC 582, NBIMCC 583 and NBIMCC 584 strains on YPD medium containing 2% glucose, which is widely used in yeast studies and provides comprehensive experiences on this enzyme (Cross and Ruis, 1978; Traczyk et al. 1985; Skoneczny and Rytka, 2000). Growth of the strains has been performed for 72 h and samples have been withdrawn during exponential and stationary phase of the culture in order to determine cell dry weight and glucose consumption (Figure 1 a, b, c). It is evident that a diauxic growth takes place as glucose is exhausted at the 12th hour of cultivation. High yield coefficient Ys is obtained (0,415 ± 0,018 - 0,423 ± 0,010). The complex nutrient medium YPD, containing organic compounds available from yeast extract and bacto-peptone caused this increase in the value of Ys in comparison to those predicted for growth of Saccharomyces cerevisiae on glucose (Walker, 1998). Samples of biomass have been processed and crude enzyme extracts have been prepared. A spectrophotometrical analysis of the total catalase activity at different hours of growth of the strains is shown in Table 1. The highest catalase activity was measured after 48 h of cultivation, which coincided, with the exponential growth phase of the cultures, after the diauxic pattern of the growth curves. This increase in catalase activity after the exhaustion of glucose in the nutrient medium could be due to the availability of different amino compounds in YPD medium, which are metabolized by fungi through hydrogen producing oxidase (Zwart et al. 1980). A correlation of the catalase activity with the accumulation of its specific substrate (H2O2) obviously takes place. An electrophoretic analysis of the crude extracts obtained from cells of Saccharomyces cerevisiae NBIMCC 583 strain of different physiological state is shown in Figure 2. After 6 hours growth, a single catalase band with Rm of 0,257 has been visualized. The next electrophoretic analyses of catalase have been performed at 12, 48 and 72 hours of cultivation, when glucose from the nutrient medium has been thoroughly consumed. An interesting fact is the appearance of a second band with Rm 0,218 at 12 and 48 hours of cultivation and its disappearance at 72 h. The catalase band, present during all cultivation period is with Rm value of 0,257. These observations are in agreement with the data published by Izawa et al. 1996 for catalase A and catalase T deficient mutants of Saccharomyces cerevisiae. They showed electrophoretic profile of the two types of catalases where the Rm value of catalase A is lower than that of catalase T. Therefore, in our experiment the protein band with Rm 0.218 corresponds to catalase A and that with Rm 0.257 – to catalase T. Catalase T we found active during the whole cultivation period. These results clearly showed that the active catalase A is synthesized after complete utilization of glucose, when catabolite repression disappeared (Cross and Ruis, 1978). During stationary phase of the culture (72 h) when catalase A was not observed, obviously a degradation of peroxisomes and lysis of the proteins took place. The above mentioned two catalase enzymes act in the cells of Saccharomyces cerevisiae, decomposing H2O2 accumulated during oxidative processes in the cytosole and correlating with the Cu/Zn superoxide dismutase activity (measured after inhibition of Mn SOD) (Table 1). There is not clear evidence about such kind of mechanism placed in mitochondria and coupling with Mn SOD. The role of H2O2 scavenging enzymes in those organelles is predicted within cytochemical experiments with cells of Saccharomyces cerevisiae by Hoffman et al. 1970. They found positive DAB reaction of intracristate space of mitochondria in the late exponential phase. These results are inconclusive due to the lack of specificity of DAB staining technique. The last could be oxidized to chromophore in the presence of H2O2 by diverse hemeproteins, including mitochondrial cytochromes (Seligman et al. 1968). This nonspecificity of the above mentioned reaction is proved also by DAB staining in catalase deficient mutant Trypanosoma cruzi (Decampo et al. 1976). Thus, clear understanding for the mitochondrial H2O2 decomposing mechanism in yeast is still lacking. Investigation of mitochondrial fraction for catalase Isolated mitochondrial fractions from Saccharomyces cerevisiae strains were tested for purity using measurement of the activities of several enzymes, typically located in different cellular compartments: hexokinase and glucose-6-phosphate dehydrogenase - in cytosole, D-amino oxidase and isocitric lyase - in peroxisomes and succinate dehydrogenase and Mn superoxide dismutase – in mitochondria. Results are presented in Table 2. Using these marker enzymes the purity of mitochondrial fraction was displayed, as the enzyme activities characteristic for the cytosole and the peroxisomes were not detected in it. The values found for the activity of succinate dehydrogenase and Mn superoxide dismutase were typical for the mitochondrial fraction, which proved that it is a pure one. Then, the specific activities of catalase and peroxidase enzymes were determined. They are 5.3 - 6.8 D E/min/mg protein for the catalase and 0.16 - 0.18 DE/min/mg protein for the peroxidase (Table 2). Further the mitochondrial fractions and crude extracts obtained from strains 582, 583 and 584 were subjected to 10% acrylamide gel electrophoresis. The results obtained are presented in Figure 3 a, b, c. All mitochondrial fractions contained a single band specifically stained for catalase. At the same time the visualization of the peroxidase activity was not successful, probably due to very low activity of the protein (Figure 4). The low peroxidase activity, measured spectrophotometrically, could be due to some kind of peroxidase activity of the catalase, as it has been shown for Candida boidinii (Ueda et al. 1990). The results obtained for the mitochondrial catalase in the three strains clearly indicated that a catalase enzyme located in the mitochondria of Saccharomyces cerevisiae exists. The Rm value of the enzyme pattern is 0.239 for the three strains. From the data presented in Figure 3, it is evident that for all strains studied, the electrophoretic pattern of the mitochondrial catalase is different from the known profile for these enzymes in Saccharomyces cerevisiae (Traczyk et al. 1985; Izawa et al. 1996). Obviously the concentrations of this protein in the total cell-free extract, where catalase A and T has been found, is very low and its visualization becomes possible after isolation of pure mitochondrial fraction. Its availability in three different strains of Saccharomyces cerevisiae indicated that it is their common feature. Evidence for correlation of mitochondrial catalase with Mn superoxide dismutase activity It is well known that catalase and Cu/Zn superoxide dismutase activities correlate during cultivation of many yeasts on different substrates and conditions (Kujumdzieva-Savova et al. 1991; Costa et al. 1993). It is worthwhile to study the activity of mitochondrial catalase, Mn and intercristate Cu/Zn superoxide dismutase enzymes in correlation with growth. These investigations were performed with the three Saccharomyces cerevisiae strains 582, 583 and 584 using the mitochondrial fraction obtained from different hours of cultivation. The dynamics of above mentioned enzyme activities are presented in Figure 5 a, b, c. These data clearly showed that the specific activities of catalase and Mn superoxide dismutase enzymes correlated with the growth and their maximum has been measured at 48 h of cultivation. The activity of Cu/Zn mitochondrial superoxide dismutase appears to be a constant one (3.5 + 0.02 ÷ 5.0 + 0.01 U/mg) during the whole period of cultivation. These results suggested that the role of mitochondrial catalase is coupled mainly with the function of manganese superoxide dismutase for detoxification of mitochondria from reactive oxygen species, generated during respiratory processes within these organelles. The presence of low peroxidase activity indicated that it also takes part in this reaction of the cells as an additional mechanism. Evidently enzymes catalase and superoxide dismutase play the basic detoxifying function in the mitochondria. High activity of antioxidant enzymes found during cultivation on YPD medium in spite of the presence of glucose as a carbon source indicated that a respiratory metabolism also takes place, because of the formation of ethanol (Walker, 1998). It is well known that Saccharomyces cerevisiae utilize glucose in aerobic conditions mainly fermentatively (Kappeli, 1986), but the diauxic growth and high values of Ys showed that utilization of additional growth substrates likely occurred. Determination of charge and molecular weight of Saccharomyces cerevisiae mitochondrial catalase The native PAGE method developed by Hedrick and Smith, 1968 provides site estimation for proteins analyzed on gels of successive high acrylamide concentration. As it has been mentioned above, the molecular weight of catalase T is between 225 - 250 kD, and for catalase A - considerably lower 170 - 190 kD. The electrophoretic pattern shows abnormal mobility of both cytosolic catalase enzymes. Applying the method of Hedrick and Smith, 1968, investigation of their behavior in native PAGE with different concentrations of acrylamide was performed (Figure 6). The results obtained (Figure 6 and Figure 7) suggest that both enzymes are charge isomers which explains why catalase A, although with lower molecular weight possess lower Rm value than catalase T. Evaluating the plot of Rm, the mitochondrial enzyme could be also considered as charge isomer. For estimation the molecular weight of the mitochondrial enzyme, comparison of its relative mobility with the ones of known proteins was done (Figure 8). Plotting the HRm against molecular weight values of the markers allowed drawing an equation, describing the electrophoretic profile (Figure 9). On this basis we could calculate the MW of the mt enzyme, which is approximately 240 000 Da. These data open the question about the origin of this new catalase protein, located in the mitochondria and its relationship to catalases A and T. References
Note: EJB Electronic Journal of Biotechnology is not responsible if on-line references cited on manuscripts are not available any more after the date of publication. Supported by UNESCO / MIRCEN network © 2002 by Universidad Católica de Valparaíso -- Chile The following images related to this document are available:Photo images[ej02011f1.jpg] [ej02011t1.jpg] [ej02011f9.jpg] [ej02011f4.jpg] [ej02011f2.jpg] [ej02011t2.jpg] [ej02011f8.jpg] [ej02011f7.jpg] [ej02011f3.jpg] [ej02011f6.jpg] [ej02011f5.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}