 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Electronic Journal of Biotechnology, Vol. 5, No. 1, April, 2002 Application of rice (Oryza sativa L.) suspension culture in studying senescence in vitro (II). Changes in DNA integrity Ramin Hosseini *#1 , Bernard J. Mulligan §2 1Plant
Research Group, YAPO School of Biological Sciences, University of Nottingham,
University Park Nottingham NG7 2RD United Kingdom E-mail: Raminh_2001@Yahoo.com Financial support: Ministry of Science, Research and Technology of Iran. Received
June 12, 2001 Code Number: ej02013 Abstract Changes in the DNA content and organisation of senescing rice cell cultures (Taipei 309) were studied, using PCR and Southern blot analyses. A mitochondrial gene (coxII), a plastid gene (psaA) and a nuclear DNA maker (RG64) were analysed. The amplification of mitochondrial (mt), plastid and nuclear DNA produced the expected fragments, indicating that there were still some intact organelles and nuclei in the senescing rice cells. However, in plastid and nuclear DNA, changes in the number and size of the PCR products were observed. Southern blot analysis revealed that mt and nuclear DNA contents declined, but plastid DNA content remained relatively the same throughout the senescence. Keywords: Programmed Cell Death (PCD), rice, senescence. Article Development and differentiation in plants require that specific cells be eliminated by cell death mechanisms (Buckner et al. 1998). Programmed Cell Death (PCD) is the genetically controlled process of cell death, which occurs during development and in response to environmental cues (Greenberg, 1996). PCD has been well characterized in animals and is a genetically controlled process that facilitates the disassembly of the dying cell (Buckner et al. 1998). Animal cells undergoing PCD can display cytoplasmic condensation and shrinkage, blebbing of plasma membranes and nuclear membranes, chromatin condensation and marginalization, internucleosomal DNA fragmentation and phagocytosis of the dying cell. When these morphological changes are observed, the cell death process is often termed apoptosis (Wyllie et al. 1980; Kressel and Groscurth, 1994). In plants, DNA degradation has been observed during certain developmental events, such as tracheary element development in pea (Mittler and Lam, 1995) in leaf senescence in rice (Sodmergen et al. 1989; Sodmergen et al. 1991), in cell senescence in Arabidopsis (Callard et al. 1996) and during the hypersensitive response to pathogen attack (Mittler and Lam, 1995; Wang et al. 1996; Mittler and Lam, 1997). In plants, changes in chloroplast ultrastructure (Greening et al. 1982; Meier and Lichtenthaler, 1982), autophagic activity (Aubert et al. 1996), accumulation of electron-dense granules in the cytoplasm, reduction in the number of mitochondrial cisternae and disorganisation of cell wall fibrillar material (Pareek et al. 1997) have been observed during senescence, environmental stress and induced starvation. Whether plant cells undergo apoptotic PCD, as defined in animal cell systems, remains an open question. In another paper (Hosseini and Mulligan, 2002) we showed growth kinetics, viability and protein content of Taipei 309 cells in a 4-week culture period. We also revealed that how acid and zinc dependent nucleases were activated in these cells. Now, a further step has been taken to discover whether the detected nuclease activity, has any effect on genomic and/or organellar DNAs, a phenomenon known to be a criterion of apoptosis in animal systems. The results are discussed in the context of current research on PCD in plants. Materials and Methods Cell suspension cultures of the rice cv. Taipei 309 were grown in 100 ml conical flasks (Finch, 1991). One ml of settled cells was inoculated in a sterilised conical flask containing 22 ml of fresh AA2 medium (Ohira et al. 1973) and 7 ml of spent medium. The cells were grown on a rotary shaker at 120 rpm at 27°C ± 1 for 3 weeks post-subculture. Every week the cells were collected (three replicates) by sieving through 30 mm sieves and air-dried for 2 h on paper towels and stored at -20°C for future use.
DNA extraction. Total DNA from Taipei 309 seedlings and 1-, 2- and 3-week old cells was isolated (Doyle and Doyle, 1987) and its concentration was determined (Sambrook et al. 1989). Primer design for PCR reactions. Three pairs of primers (20) bases were designed to allow amplification of nuclear, plastid and mitochondrial gene sequences (one of each). Published rice sequences were inspected to determine suitable sequences as follows:
The three primers (Table 1) were synthesised through the Department of Biochemistry, Queens Medical Centre, University of Nottingham and used for PCR reactions. The length of the gene/marker and size of expected amplified fragments are given in Table 2. Also PCR products were radiolabelled, using a -[32p](dCTP; Amersham Life Science Ltd., Little Chalfont, UK ) and used as hybridisation probes. Conditions employed for PCR. Total DNA from Taipei 309 seedlings was used for the optimisation of the PCR conditions. To optimise the PCR conditions, different concentrations (0.5 mM, 1 mM, 1.5 mM and 2 mM) of MgCl2 in the reaction buffer and two annealing temperatures (45 and 50°C) were tested (see Results). After optimisation of PCR conditions, DNA samples from Taipei 309 seedlings and 1-, 2- and 3-week old Taipei 309 cell cultures were used for investigation of the integrity of coxII, psaA and RG64. The amplification of the desired DNA fragments was carried out based on the method described in Sambrook et al. 1989. Southern blot analysis. The amplified fragments of mt, plastid and nuclear DNA (see above) obtained by the PCR reactions from Taipei 309 seedlings were used as probes and Southern blot analysis of senescing cells of Taipei 309 was carried out; following the method described by Sambrook et al. 1989. Analysis and purification of the amplified DNA fragments. To analyse the PCR products, 6 m l of the reaction mixture was mixed with 1 m l of 6 x loading buffer [0.25% (w/v) bromophenol blue, 0.25% (w/v) xylene cyanol FF, 30% (v/v) glycerol] and electrophoresed on a 0.8% (w/v) agarose gel (5 x 6 cm; Sigma, type V) at 100 v for 30 min. The gel was stained with ethidium bromide (0.5 mg/l) for 30 min and the PCR products were visualised and photographed using a UV transilluminator (TM-20 model; UVP Ltd., California, USA). The amplified fragments were purified using a Prep-A-Gene kit according to the manufacturer's recommendation (Bio-Rad, Hemel Hempstead, UK).
Results
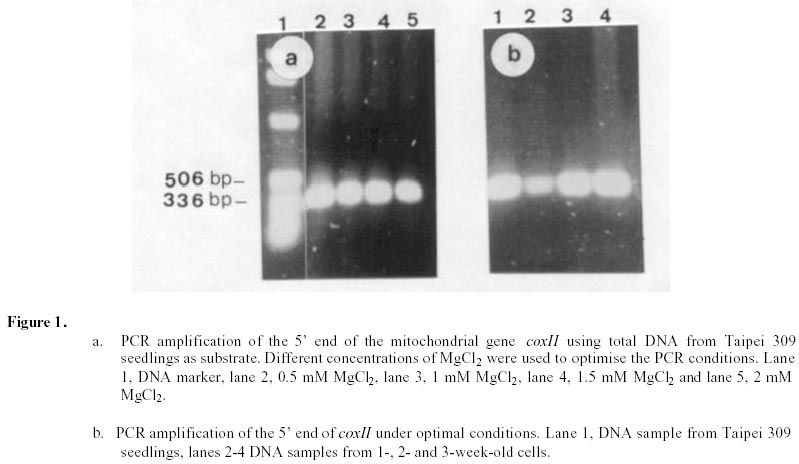
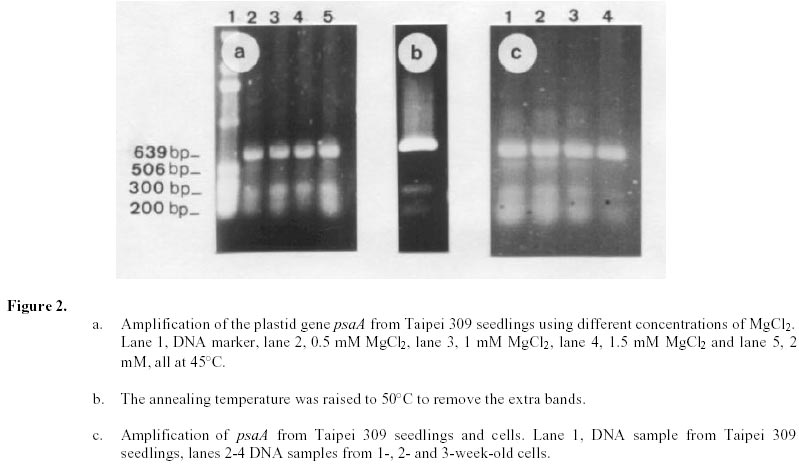
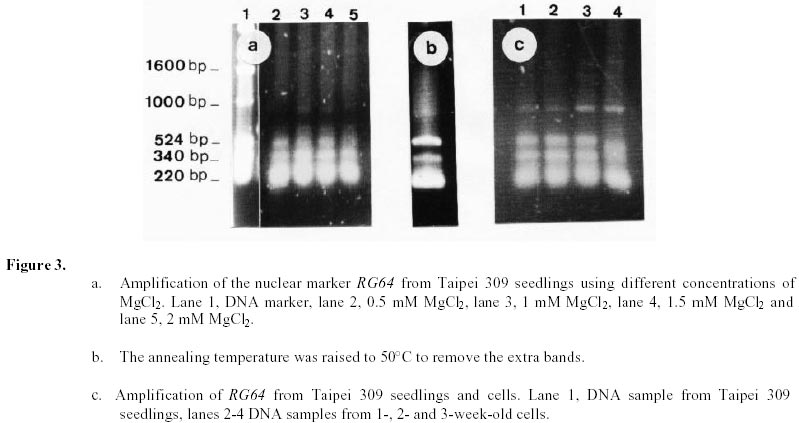
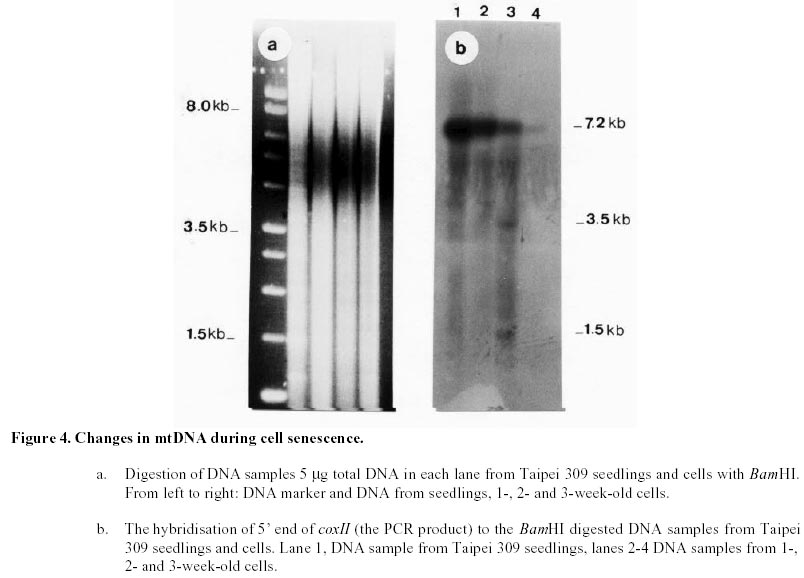
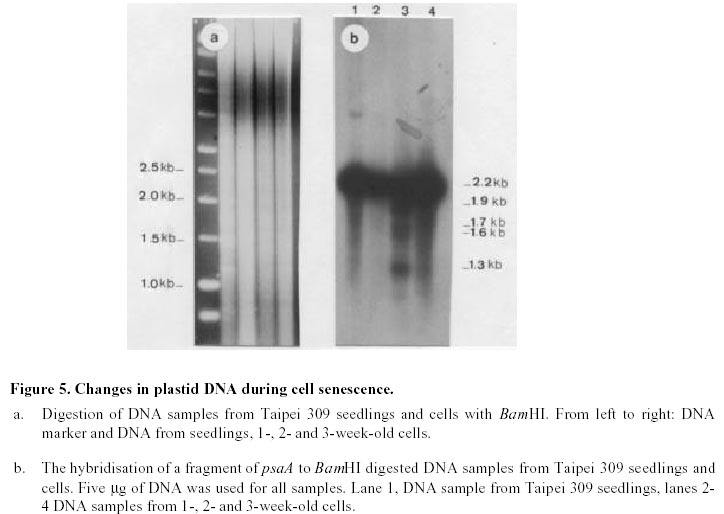
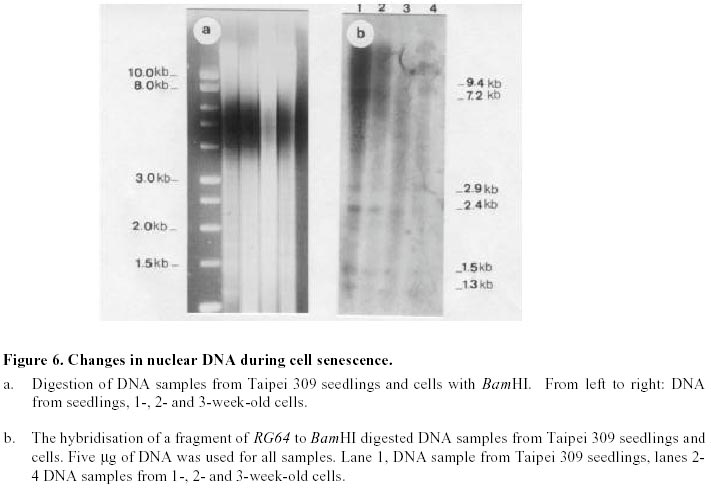
Changes in DNA integrity in senescing Taipei 309 cells, revealed by PCR The integrity of mitochondrial, chloroplast and nuclear DNA during senescence of Taipei 309 cells was investigated using PCR. The DNA from Taipei 309 seedlings was used as a control, alongside DNA samples from 1-,2- and 3-week old Taipei 309 cells. Mitochondrial DNA (coxII) PCR amplification of the 5’ end of the rice coxII gene was carried out under the conditions described previously. Different concentrations of MgCl2 were tested in the PCR. A 2mM concentration of MgCl2 slightly inhibited the amplification of the coxII product, compared to the lower concentrations tested (Figure 1a). Therefore, 0.5 mM MgCl2 was chosen as the optimum concentration. The expected fragment (336 bp) was amplified in all conditions used. In order to investigate the intactness of the mitochondrial DNA (coxII), PCR reactions were carried out using 0.05 m g of template DNA from Taipei 309 seedlings and from 1-, 2- and 3-week old cells. No differences were observed in number and size of the amplified products between samples (Figure 1b). Plastid DNA (psaA) Different concentrations of MgCl2 were tested to optimise the PCR conditions and 2 mM MgCl2 was found to give the best amplification. The expected fragment (639 bp) was amplified but two smaller and diffuse (200 and 300 bp) products were also observed (Figure 2a). An attempt to restrict amplification of the extra bands by increasing the annealing temperature to 50°C failed (Figure 2b). In order to investigate the integrity of plastid DNA in the senescing Taipei 309 cells, a PCR reaction with DNA samples from Taipei 309 seedlings, 1-, 2- and 3-week old cells was run under optimal conditions. The 639 bp was amplified in all samples (Figure 2c) but the two diffuse bands were less obvious in the PCR product from 2-week-old cells and barely detectable in the PCR product from 3-week-old cells (Figure 2c). Nuclear DNA (RG64) The optimum MgCl2 concentration was found to be 1 mM (Figure 3a). In addition to the expected product (524 bp), two smaller fragments (about 220 and 340 bp) were also observed (Figure 3a). The same products were amplified when the primer annealing temperature was increased to 50°C (Figure 3b). Changes in nuclear DNA (RG64) integrity were investigated by running a PCR reaction using DNA templates from Taipei 309 seedlings, 1-, 2- and 3-week old cells under optimal conditions. In addition to the expected product (524 bp), as observed when using DNA from cultured cells, three other products (220 bp, 340 bp, 1000 bp) were amplified in the Taipei 309 seedling sample (Figure 3c). The abundance of the 1000 bp product was greater when PCRs were carried out using DNA from 2- and 3-week-old cells compared to seedlings and 1-week-old cells. The other amplified DNA products (220 and 340 bp) from 1- and 2- week- old cells were similar in abundance to those produced from seedlings, though the 524 bp and 340 bp products were reduced in 3-week-old cells (Figure 3c). In the amplified products from 3-week-old cells, a faint extra band of about 1600 bp was also produced (Figure 3c). Southern blot analysis of mitochondrial, plastid and nuclear DNA of senescing cells of Taipei 309 Southern blot analysis of DNA from Taipei 309 cells was carried out in order to detect any changes in the organellar and/or nuclear DNA as the cells senesced. PCR amplified fragments of a mitochondrial gene, coxII, a plastid gene, psaA, and a nuclear DNA marker, RG64 were radiolabelled and used as probes (see above). Mitochondrial DNA (coxII) Total DNA samples (5 mg) from Taipei 309 seedlings, 1-, 2- and-3-week old cells were digested with BamHI (Figure 4a) and hybridised to a PCR fragment corresponding to the 5’ end of the rice mitochondrial gene, coxII (see above). In seedlings and 1-week old cells, the probe hybridised to one fragment (7.2 kb). In 2-week-old cells, however, 2 extra bands of 3.5 and 1.5 kb were also observed. The intensity of the 7.2 kb signal decreased with cell age and in 3-week-old cells, only a very weak hybridisation to the 7.2 kb fragment was observed (Figure 4b). It was clear that DNA content declined progressively in 1- to 3-week-old cells (Figure 4b). Plastid DNA (psaA) Total DNA (5 mg) from Taipei 309 seedlings, 1-, 2- and 3-week-old cells was digested with BamHI (Figure 5a) and hybridised to a PCR fragment of the rice chloroplast gene psaA. In the seedlings and cell samples there was a strong 2.2 kb signal whose intensity remained the same regardless of cell age. In addition to this band, in 2-week-old cells, four very weak signals of 1.9, 1.7, 1.6 and 1.3 kb were observed. In 3-week-old cells, the hybridisation pattern was similar to that of 2-week-old cells but the 1.9, 1.7, 1.6 and 1.3 kb signals were unclear (Figure 5b). Nuclear DNA (RG64) For nuclear DNA analysis, total DNA (5 m g) was digested with BamHI (Figure 6a), and hybridised to a PCR fragment of RG64, a rice nuclear DNA marker. All samples from seedlings and cells showed 6 signals, with sizes of 9.4, 7.2, 2.9, 2.4, 1.5 and 1.3 kb. The strongest signals were obtained with seedling DNA but the signals became progressively weaker in 1-, 2- and 3-week-old cells (Figure 6b). Discussion Changes in DNA integrity of senescing rice cells Senescence is a controlled sequence of events comprising the final stage of development (Greening et al. 1982; Guiamet and Giannibelli, 1994; Smart, 1994). The existence of senescence-related genes provides the most direct evidence for senescence being controlled by gene expression (Thomas and Howarth, 2000) as this has been recently confirmed by Hinderhofer and Zentgraf, 2001. DNA degradation is one of the events occurring during senescence (Sodmergen et al. 1989; Sodmergen et al. 1991). DNA degradation has been reported to occur during other plant developmental stages (Blank and McKeon, 1989; Mittler and Lam, 1995; Callard et al. 1996; Greenberg, 1996; Ryerson and Heath, 1996; Wang et al. 1996). In the present study, in order to find out any changes in DNA related to cell senescence, the integrity of nuclear, mt and plastid DNA of Taipei 309 cells was investigated by PCR and Southern blot analysis. mtDNA As expected, use of the coxII primers in PCRs, with total DNA from rice seedlings as substrate, produced a 336 bp amplified DNA fragment. The same sized fragment was produced when total DNA from 1-3 week old suspension cultures was employed as substrate. This result suggested that some intact mitochondrial DNA was still present even in 3-week-old cultures. Under the PCR conditions employed, it was not possible to conclude whether mitochondrial DNA (per cell or per g.f.wt. cells) decreased from week 1 to week 3, neither was it possible on the basis of the PCR data alone to assess the integrity of the mtDNA. However, the Southern hybridisation data suggested that the proportion of total DNA represented by the mt genome decreased with cell age and at the same time changes in mt DNA occurred. Aubert et al. 1996 suggested that in plants, signal(s) giving rise to the autophagic process originate(s) from the decline of supply of respiratory substrates to the mitochondria. It seems that in plant senescence, the role of mitochondrial DNA alterations has not been fully investigated, even though the mitochondria have been found intact until the late stages of senescence (Smart, 1994). The data presented here suggested that mtDNA alterations were occurring in the rice cells as they senesced. Plastid DNA psaA and psaB genes encode for the two polypeptides of P700 chlorophyll a protein complex of photosystem I reaction centre. These two genes are located adjacent to each other in the large single copy region of the circular plastid genome in higher plants (Chen et al. 1992). Primers for plastid DNA (psaA) showed several amplified bands in PCR, whereas since psaA is a single copy sequence (Hiratsuka et al. 1989), only one band was expected. Increasing the annealing temperature to 50°C failed to solve the problem of the extra bands. It may suggest that there are homologous sequences to the investigated sequence (psaA) in the rice genome. The expected psaA band was amplified in seedlings and cell samples. In seedlings, 1- and 2-week old cells the two diffuse bands (200 and 300 bp) were present but in the PCR product from 3-week-old cultures, the intensity of the two extra bands reduced. Thus, as judged by PCR, plastid DNA showed some differences in 2- and 3-week-old cells compared to the seedlings and 1-week-old cells. These findings may suggest that subtle changes had taken place in plastid DNA of the senescing rice cells. Interestingly, however, Southern blots showed that the abundance of plastid DNA was only slightly affected by cell age. There are contradictory reports about chloroplast integrity during senescence. Wittenbach et al. 1982 reported that chloroplasts could be isolated from protoplasts prepared from dark-induced senescent wheat leaves from which up to 80% of chlorophyll and protein had been lost. Consistent with the results presented here, however, Jones and Dangl, 1996 mentioned that plastids appear to be the last organelles to be degraded. Greening et al. 1982 reported the loss of plastid integrity, while Sodmergen et al. 1989 and Sodmergen et al. 1991 reported a decline in cpDNA at early stages of senescence. However, in this research, it was observed that in rice cells, the quantity of plastid DNA remained the same throughout the process of senescence; even though qualitative alterations of plastid DNA were observed. Bate et al. 1990, reported that during senescence in bean plants, D-1 the product of psaA, a PSII (photosystem II) protein, continued to be actively synthesised despite a 9-fold decrease in RNA synthesis and the reduced synthesis of all major thylakoid proteins. They also found that in senescent bean, the synthesis of psaA RNA was unaltered during senescence. In the present study, Southern blot analysis of psaA in senescing Taipei309 cells revealed that this gene remained intact up to very late stages of senescence. Significant transcript levels of psaA and psaB have also been observed in dark grown barley seedlings (Kreus et al. 1986). Nuclear DNA When the amplification of RG64 from seedlings and cells was carried out, several extra PCR products were detected in addition to the expected product (524 bp). However, in 3-week-old cells the 524 bp band was reduced in abundance and became diffuse but one of the extra bands (1000 bp product) intensified and another extra band about 1.6 kb was produced. Nuclear DNA samples showed the same hybridisation pattern to the RG64 probe but a decrease in DNA quantity with cell age was observed. Guarente, 1996 suggests that changes in chromosomal structure or function may be a key determinant of ageing. In plants, nuclei remain intact up to the late stages of senescence (Smart, 1994). It has been suggested that nuclear genes control the degradation of chloroplasts (Guiamet and Giannibelli, 1994). Concluding Remarks The PCR assay revealed no alteration to mtDNA in ageing rice cells, whereas Southern blot showed 2 extra signals in 2-week-old cells and a progressive decrease in mtDNA content. Plastid DNA had undergone some rearrangements, which was shown by both PCR and Southern blot analysis, with no decrease in its quantity. Thus, qualitative and quantitative changes were detected in nuclear DNA by PCR and Southern blot analysis, respectively. Nuclear and mtDNA content of the senescing rice cells declined. However, plastid DNA content showed subtle changes. The decrease in DNA content may imply an increase in nuclease activity in senescing cells. DNA degradation, a feature of animal cell death, apoptosis, was observed in senescing rice cells. However, the changes in DNA content and integrity should be more fully investigated using a number of genes. Following the investigation of changes at DNA level, discovering changes at transcription and translation levels would also be beneficial. References
Note: EJB Electronic Journal of Biotechnology is not responsible if on-line references cited on manuscripts are not available any more after the date of publication. Supported by UNESCO / MIRCEN network © 2002 by Universidad Católica de Valparaíso -- Chile The following images related to this document are available:Photo images[ej02013f4.jpg] [ej02013t1.jpg] [ej02013f5.jpg] [ej02013f1.jpg] [ej02013f3.jpg] [ej02013f2.jpg] [ej02013f6.jpg] [ej02013t2.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}