 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Electronic Journal of Biotechnology, Vol. 5, No. 1, April, 2002 REVIEW ARTICLE Plant protease inhibitors in control of phytophagous insects Paulraj K. Lawrence*1 and Kripa Ram Koundal 2 1Department
of Biological Sciences, Texas Tech University, Box No 43131 Lubbock, TX 79409-3131,
USA Tel: 806 742 2740 Fax: 806 742 2963 E-mail: p.k.lawrence@ttu.edu Received
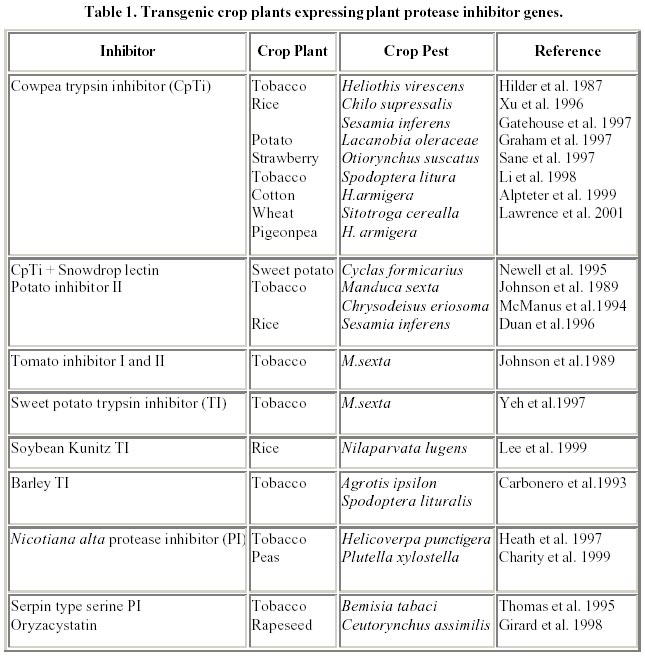
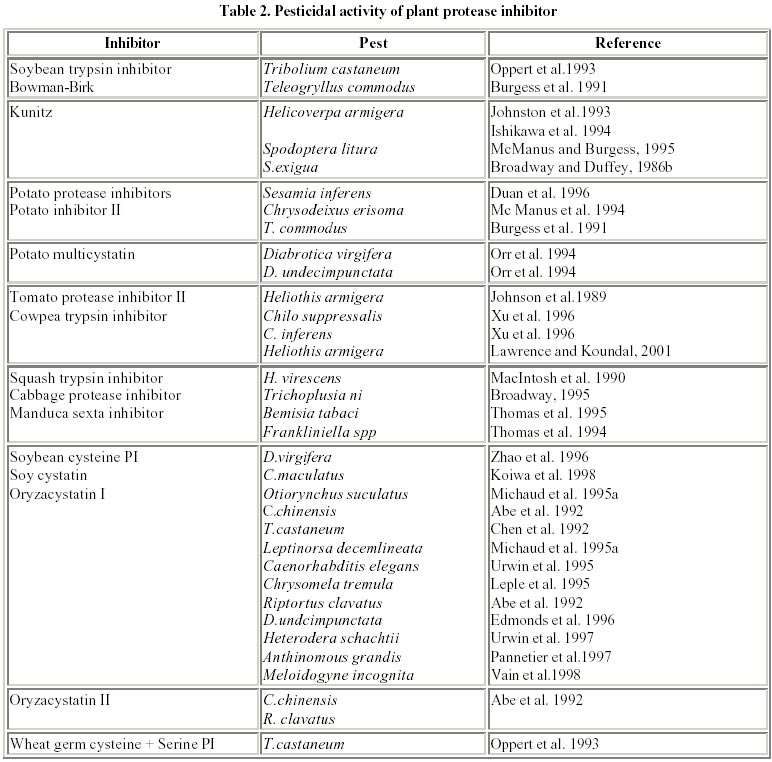
August 20, 2001 Code Number: ej02015 Abstract Plant proteinase inhibitors (PIs) have been well established to play a potent defensive role against predators and pathogens. Although diverse endogenous functions for these proteins has been proposed, ranging from regulators of endogenous proteinases to act as storage proteins, evidence for many of these roles is partial, or confined to isolated examples. On the other hand, many PIs have been shown to act as defensive compounds against pests by direct assay or by expression in transgenic crop plants, and a body of evidence for their role in plant defense has been accumulated consistently. The role and mechanism of action for most of these inhibitors are being studied in detail and their respective genes isolated. These genes have been used for the construction of transgenic crop plants to be incorporated in integrated pest management programmes. This article describes the classes of protease inhibitors, their regulation and genes used to construct transgenic plants against phytophagous insects. Keywords: crop pests, protease inhibitors, transgenic plants. Article World wide crop losses without the use of pesticides and other non-chemical control strategies is estimated to be about 70% of crop production, amounting to US $ 400 billion. The world wide pre-harvest losses due to insect pests, despite, the use of insecticides is 15% of total production representing over US $ 100 billion (Krattiger, 1997). The annual cost of insect control itself amounts to US $ 8 billion, thus warranting urgent economical control measures. Many of the crop varieties developed in the past 30 years were high yielders, but had poor storage characteristics (Kerin, 1994). Insect pests are capable of evolving to biotypes that can adapt to new situations, for instance, they overcome the effect of toxic materials or bypass natural or artificial plant resistance, which further confounds the problem (Roush and McKenzie, 1987). Under these circumstances, provision of food to the rapidly expanding population has always been a challenge facing mankind. This problem is more acute in the tropics and sub-tropics, where the climate provides a highly condusive environment for a wide range of insects and necessitates massive efforts to suppress the population densities of different pests in order to achieve an adequate supply of food. In developing countries, the problem of competition from insect pests is further complicated with a rapid annual increase in the human population (2.5-3.0 percentage) in comparison to a 1.0 percent increase in food production. In order to feed the ever expanding population, crop protection plays a vital and integral role in the modern day agricultural production to minimise yield losses. Currently, the crop protection practice in such agricultural systems relies exclusively on the use of agrochemicals, although a few specific cases do exist where inherent varietal resistance and biological control have been successfully employed. The exclusive use of chemical pesticides not only results in rapid build-up of resistance to such compounds, but their non-selectivity affects the balance between pests and natural predators, and is generally in favour of pests (Metcalf, 1986). Therefore, an integrated pest management (IPM) programme, comprising a combination of practices including the judicious use of pesticides, crop rotation, field sanitation and above all exploitation of inherently resistant plant varieties would provide the best option (Meiners and Elden, 1978). The last option includes the use of transgenic crops, expressing foreign insecticidal genes which could make a significant contribution to sustainable agriculture and thus could be an important component of IPM (Boulter, 1993). The production of transgenic crops has seen rapid advances during the last decade with the commercial introduction of Bt transgenics, but the major concern with these crops has been the development of resistance by pest and public acceptability. Hence, there has been a need to discover new effective plant genes which would offer resistance/protection against these pests. Protease inhibitors (PIs) are one of the prime candidates with highly proven inhibitory activity against insect pests and also known to improve the nutritional quality of food. Plant protease inhibitors The possible role of protease inhibitors (PIs) in plant protection was investigated as early as 1947 when, Mickel and Standish observed that the larvae of certain insects were unable to develop normally on soybean products. Subsequently the trypsin inhibitors present in soybean were shown to be toxic to the larvae of flour beetle, Tribolium confusum (Lipke et al. 1954). Following these early studies, there have been many examples of protease inhibitors active against certain insect species, both in in vitro assays against insect gut proteases (Pannetier et al. 1997; Koiwa et al, 1998) and in in vivo artificial diet bioassays (Urwin et al. 1997; Vain et al. 1998). The term “protease” includes both “endopeptidases” and “exopeptidases” whereas, the term “proteinase” is used to describe only “endopeptidases” (Ryan, 1990). Several non-homologous families of proteinase inhibitors are recognized among the animal, microorganisms and plant kingdom. Majority of proteinase inhibitors studied in plant kingdom originates from three main families namely leguminosae, solanaceae and gramineae (Richardson, 1991). These protease inhibitor genes have practical advantages over genes encoding for complex pathways i.e. by transferring single defensive gene from one plant species to another and expressing them from their own wound inducible or constitutive promoters thereby imparting resistance against insect pests (Boulter, 1993). This was first demonstrated by Hilder et al. 1987 by transferring trypsin inhibitor gene from Vigna unguiculata to tobacco, which conferred resistance to wide range of insect pests including lepidopterans, such as Heliothis and Spodoptera, coleopterans such as Diabrotica, Anthonomnous and orthoptera such as Locusts. Further, there is no evidence that it had toxic or deleterious effects on mammals. Many of these protease inhibitors are rich in cysteine and lysine, contributing to better and enhanced nutritional quality (Ryan, 1989). Protease inhibitors also exhibit a very broad spectrum of activity including suppression of pathogenic nematodes like Globodera tabaccum, G. pallida, and Meloidogyne incognita by CpTi (Williamson and Hussey, 1996), inhibition of spore germination and mycelium growth of Alternaria alternata by buckwheat trypsin/chymotrypsin (Dunaevskii et al. 1997) and cysteine PIs from pearl millet inhibit growth of many pathogenic fungi including Trichoderma reesei (Joshi et al. 1998). These advantages make protease inhibitors an ideal choice to be used in developing transgenic crops resistant to insect pests. Further, transformation of plant genomes with PI-encoding cDNA clones appears attractive not only for the control of plant pests and pathogens, but also as a means to produce PIs, useful in alternative systems and the use of plants as factories for the production of heterologous proteins (Sardana et al. 1998). These inhibitor families that have been found are specific for each of the four mechanistic classes of proteolytic enzymes, and based on the active amino acid in their “reaction center” (Koiwa et al. 1997), are classified as serine, cysteine, aspartic and metallo-proteases. Serine proteinase inhibitors The role of serine PIs as defensive compounds against predators is particularly well established, since the major proteinases present in plants, used for processes such as protein mobilization in storage tissues, contain a cysteine residue as the catalytically active nucleophile in the enzyme active site. Serine proteinases are not used by plants in processes involving large scale protein digestion, and hence the presence of significant quantities of inhibitors with specificity towards these enzymes in plants cannot be used for the purposes of regulating endogenous proteinase activity (Reeck et al. 1997). In contrast, a major role for serine PIs in animals is to block the activity of endogenous proteinases in tissues where this activity would be harmful, as in case of pancreatic trypsin inhibitors found in mammals. The serine class of proteinases such as trypsin, chymotrypsin and elastase, which belong to a common protein superfamily, are responsible for the initial digestion of proteins in the gut of most higher animals (GarciaOlmedo et al. 1987). In vivo they are used to cleave long, essentially intact polypeptide chains into short peptides which are then acted upon by exopeptidases to generate amino acids, the end products of protein digestion. These three types of digestive serine proteinases are distinguished based on their specificity, trypsin specifically cleaving the C-terminal to residues carrying a basic side chain (Lys, Arg), chymotrypsin showing a preference for cleaving C-terminal to residues carrying a large hydrophobic side chain (Phe, Tyr, Leu), and elastase showing a preference for cleaving C-terminal to residues carrying a small neutral side chain (Ala, Gly) (Ryan, 1990). Inhibitors of these serine proteinases have been described in many plant species, and are universal throughout the plant kingdom, with trypsin inhibitors being the most common type. At least, part of this bias can be accounted for by the fact that (mammalian) trypsin is readily available and is the easiest of all the proteinases to assay using synthetic substrates, and hence is used in screening procedures. Because of these reasons the members of the serine class of proteinases have been the subject of intense research than any other class of proteinase inhibitors. Such studies have provided a basic understanding of the mechanism of action (Huber and Carrell, 1989) that applies to most serine proteinase inhibitor families and probably to the cysteine and aspartyl proteinase inhibitor families as well. All serine inhibitor families from plants are competitive inhibitors and all of them inhibit proteinases with a similar standard mechanism (Laskowski and Kato, 1980). Serine proteinases have been identified in extracts from the digestive tracts of insects from many families, particularly those of lepidoptera (Houseman et al. 1989) and many of these enzymes are inhibited by proteinase inhibitors. The order lepidoptera, which includes a number of crop pests, the pH optima of the guts are in the alkaline range of 9-11 (Applebaum, 1985) where, serine proteinases and metallo-exopeptidases are most active. Additionally, serine proteinase inhibitors have anti-nutritional effects against several lepidopteran insect species (Shulke and Murdock, 1983; Applebaum, 1985). Purified Bowman-Birk trypsin inhibitor (Brovosky, 1986) at 5% of the diet inhibited growth of these larvae but SBTI (Kunitz, 1945), another inhibitor of bovine trypsin, was less effective when fed at the same levels. Broadway and Duffey (1986a) compared the effects of purified SBTI and potato inhibitor II (an inhibitor of both trypsin and chymotrypsin) on the growth and digestive physiology of larvae of Heliothis zea and Spodoptera exigua and demonstrated that growth of larvae was inhibited at levels of 10% of the proteins in their diet. Trypsin inhibitors at 10% of the diet were toxic to larvae of the Callosobruchus maculatus (Gatehouse and Boulter, 1983) and Manduca sexta (Shulke and Murdock, 1983). Recent X-ray crystallography structure of winged bean, Psophocarpus tetragonolobus Kunitz-type double headed alpha-chymotrypsin shows 12 anti-parallel beta strands joined in a form of beta trefoil with two reactive site regions (Asn 38-Leu 43 and Gln 63-Phe 68) at the external loops (Ravichandaran et al. 1999; Mukhopadhyay, 2000). Structural analysis of the Indian finger millet (Eleusine coracana) bifunctional inhibitor of alpha-amylase/trypsin with 122 amino acids has shown five disulphide bridges and a trypsin binding loop (Gourinath et al. 2000). These structural analysis would greatly help in “enzyme engineering” of the native PIs to a potent form, against the target pest species than the native PIs. Cysteine proteinase inhibitors Isolation of the midgut proteinases from the larvae of cowpea weevil, C. maculatus (Kitch and Murdock, 1986; Campos et al. 1989) and bruchid Zabrotes subfaceatus (Lemos et al. 1987) confirmed the presence of cysteine mechanistic class of proteinase inhibitors. Similar proteinases have been isolated from midguts of the flour beetle Tribolium castaneum, Mexican beetle Epilachna varivestis (Murdock et al. 1987) and the bean weevil Ascanthoscelides obtectus (Wieman and Nielsen, 1988). Cysteine proteinases isolated from insect larvae are inhibited by both synthetic and naturally occurring cysteine proteinase inhibitors (Wolfson and Murdock, 1987). In a study of the proteinases, from the midguts of several members of the order coleopteran, 10 of 11 beetle species representing 11 different families, had gut proteinases that were inhibited by p-chloromercuribenzene sulfonic acid (PCMBS), a potent sulphydryl reagent (Murdock et al. 1988) indicating that the proteinases were of the cysteine mechanistic class. The optimum activity of cysteine proteinases is usually in the pH range of 5-7, which is the pH range of the insect gut that use cysteine proteinases (Murdock et al. 1987). Another puzzling aspect of studies with C. maculatus is the apparent effects of certain members of Bowman-Birk trypsin inhibitor family on the growth and development of these larvae. Although cysteine proteinase is primarily responsible for protein digestion in C. maculatus, it is not clear, how the cowpea and soybean Bowman-Birk inhibitors are exert their anti-nutritional effects on this organism. Advances in enzymology has revealed the existence of a variety of cysteine proteinases resulting in their classification into several families namely papain, calpin and asparagines specific processing enzyme (Turk and Bode, 1991). Cystanins have also been characterized from potato (Waldron et al. 1993), ragweed (Rogers et al. 1993), cowpea (Fernandes et al. 1993) papaya (Song et al. 1995) and avacado (Kimura et al. 1995). The rice cysteine proteinase inhibitors are the most studied of all the cysteine PIs which is proteinaceous in nature (Abe and Arai, 1985) and highly heat stable (Abe et al. 1987). Recent three dimensional structure analysis of oryzacystatin OC-I by Tanokura’s group (Nagata et al. 2000), using NMR has showed a well defined main body consisting of amino acids from Glu 13 - Asp 97 and an alpha helix with five stranded anti parallel beta-sheet, while the N terminus (Ser 2-Val 12) and C terminus (Ala 98-Ala 102) are less defined. Further, analysis has demonstrated OC-I to be similar to chicken cystatin which belongs to type-2 animal cystatin. Another rice cystatin named as OC-II, with a putative target binding motif gln-x-val-x-gly shares similar motif with OC-I but has a different inhibition constant (Ki) value (Arai et al. 1991; Kondo et al. 1991). Aspartic and metallo-proteinase inhibitors Knowledge on the role of aspartic proteinases in insect digestion is limited than that of cysteine proteinases. In species of six families of the order hemiptera, aspartic proteinases (cathepsin D-like proteinases) were found along with cysteine proteinases (Houseman and Downe, 1983). The low pH of midguts of many members of coleoptera and hemiptera provides more favourable environments for aspartic proteinases (pH optima ~ 3-5) than the high pH of most insect guts (pH optima ~ 8-11) (Houseman et al. 1987) where the aspartic and cysteine proteinases would not be active. No aspartic proteinases have been isolated from coleoptera but Wolfson and Murdock 1987 demonstrated that pepstatin, a powerful and specific inhibitor of aspartyl proteinases, strongly inhibited proteolysis of the midgut enzymes of Colorado potato beetle, Leptinotarsa decemlineata indicating that an aspartic proteinase was present in the midgut extracts. Potato tubers possess an aspartic proteinase inhibitor, cathepsin D (Mares et al. 1989) that shares considerable amino acid sequence identity with the trypsin inhibitor SBTI from soybeans. Plants have also evolved at least two families of metallo-proteinase inhibitors, the metallo-carboxypeptidase inhibitor family in potato (Rancour and Ryan, 1968), and tomato plants (Graham and Ryan, 1981) and a cathepsin D inhibitor family in potatoes (Keilova and Tomasek, 1976). The cathepsin D inhibitor (27 kDa) is unusual as it inhibits trypsin and chymotrypsin as well as cathepsin D, but does not inhibit aspartyl proteases such as pepsin, rennin or cathepsin E. The inhibitors of the metallo-carboxypeptidase from tissue of tomato and potato are polypeptides (4 kDa) that strongly and competitively inhibit a broad spectrum of carboxypeptidases from both animals and microorganisms, but not the serine carboxypeptidases from yeast and plants (Havkioja and Neuvonen, 1985). The inhibitor is found in tissues of potato tubers where it accumulates during tuber development along with potato inhibitor I and II families of serine proteinase inhibitor. The inhibitor also accumulates in potato leaf tissues along with inhibitor I and II proteins in response to wounding (Graham and Ryan, 1981; Hollander-Czytko et al. 1985). Thus, the inhibitors accumulated in the wounded leaf tissues of potato have the capacity to inhibit all the five major digestive enzymes i.e. trypsin, chymotrypsin, elastase, carboxypeptidase A and carboxypeptidase B of higher animals and many insects (Hollander-Czytko et al. 1985). Aspartic PIs have been recently been isolated from sunflower (Park et al. 2000), barley (Kervinen et al. 1999) and cardoon (Cyanara cardunculus) flowers named as cardosin A (Frazao et al. 1999). The detailed structural analysis of prophytepsin, a zymogen of barley aspartic proteinase shows a pepsin like bilobe and a plant specific domain. The N terminal has 13 amino acids necessary for inactivation of the mature phytepsin (Kervinen et al. 1999), and the aspartic PI cardosin A from cardoon shows regions of glycolylations at Asn-67 and Asn 257. The Arg-Gly-Asp sequences recogonizes the cardosin receptor which is found in a loop between two-beta strands on the molecular surface (Frazao et al. 1999). Mechanism of toxicity The mechanism of action of these proteinase inhibitors has been a subject of intense investigation (Barrett, 1986; MacPhalen and James, 1987; Greenblatt et al. 1989). Knowledge on mechanisms of protease action and their regulation in vitro, and in vivo, in animals, plants, microorganisms and more recently in viruses have contributed to many practical applications for inhibitor proteins in medicine and agriculture. Baker et al. 1984 showed that the secretion of proteases in insect guts depends upon the midgut protein content rather than the food volume. The secretion of proteases has been attributed to two mechanisms, involving either a direct effect of food components (proteins) on the midgut epithelial cells, or a hormonal effect triggered by food consumption (Applebaum, 1985). Models for the synthesis and release of proteolytic enzymes in the midguts of insects proposed by Birk and Applebaum, 1960, Brovosky, 1986 reveal that ingested food proteins trigger the synthesis and release of enzymes from the posterior midgut epithelial cells. The enzymes are released from membrane associated forms and sequestered in vesicles that are in turn associated with the cytoskeleton. The peptidases are secreted into the ectoperitrophic space between the epithelium, as a particulate complex (Eguchi et al. 1982), from where the proteases move transversely into the lumen of the gut, where the food proteins are degraded. PIs inhibit the protease activity of these enzymes and reduce the quantity of proteins that can be digested, and also cause hyper-production of the digestive enzymes which enhances the loss of sulfur amino acids (Shulke and Murdock, 1983) as a result of which, the insects become weak with stunted growth and ultimately die. The digestive proteolytic enzymes in the different orders of commercially important insect pests belong to one of the major classes of proteinases predominantly. Coleopteran and hemipteran species tend to utilize cysteine proteinases (Murdock et al. 1987) while lepidopteran, hymenopteran, orthopteran and dipteran species mainly use serine proteinases (Ryan, 1990; Wolfson and Murdock, 1990). Examples from both of these classes of proteinases have been shown to be inhibited by their cognate proregions (Taylor et al. 1995). The effect of class specific inhibitors on the pest digestive enzymes is not always a simple inhibition of proteolytic activity, but recent studies have indicated the reverse may happen. It would appear that there are often two populations of digestive enzymes in target pests, those that are susceptible to inhibition and those that are resistant (Michaud et al. 1996; Bown et al. 1997), and some insects respond to ingestion of plant PIs such as soybean trypsin inhibitor (Broadway and Duffey, 1986b) and oryzacystatin (Michaud et al. 1996) by hyper-producing inhibitor-resistant enzymes. The mechanism of binding of the plant protease inhibitors to the insect proteases appear to be similar with all the four classes of inhibitors. The inhibitor binds to the active site on the enzyme to form a complex with a very low dissociation constant (107 to 1014 M at neutral pH values), thus effectively blocking the active site. A binding loop on the inhibitor, usually "locked" into conformation by a disulphide bond, projects from the surface of the molecule and contains a peptide bond (reactive site) cleavable by the enzyme (Terra et al. 1996; Walker et al. 1998). This peptide bond may be cleaved in the enzyme inhibitor complex, but cleavage does not affect the interaction, so that a hydrolyzed inhibitor molecule is bound similar to an unhydrolyzed one. The inhibitor thus directly mimics a normal substrate for the enzyme, but does not allow the normal enzyme mechanism of peptide bond cleavage to proceed to completion ie., dissociation of the product (Walker et al. 1998). It would also appear that insect digestive trypsins do not fall into the classification of peptidase hydrolases, as defined by inhibition spectra. It has been shown, notably, that the trypsin like digestive proteases of several lepidopteran species are inhibited by (l-3-trans carboxiran-2-carbonyl)-l-leu-agmatin (E-64) (Lee and Anstee, 1995; Novillo et al. 1997) an inhibitor generally considered to be specific to cysteine proteinases (Dunn, 1989). Thus, true interactions will become clear only when we have protein crystals and X-ray diffraction data for the structure of insect enzyme/inhibitor complexes. Further, specificity of the inhibitor enzyme interaction is primarily determined by the specificity of proteolysis determined by the enzyme (Blancolabra et al. 1996). Regulation of proteinase inhibitors Plant proteinase inhibitor proteins that are known to accumulate in response to wounding have been well characterized. Earlier research on tomato inhibitors has shown that the protease inhibitor initiation factor (PIIF), triggered by wounding/injury switches on the cascade of events leading to the synthesis of these inhibitor proteins (Melville and Ryan, 1972; Bryant et al. 1976), and the newly synthesized PIs are primarily cytosolic (Hobday et al. 1973; Meige et al. 1976). The current evidence suggests that the production of the inhibitors occurs via. the octadecanoid (OD) pathway, which catalyzes the break down of linolenic acid and the formation of jasmonic acid (JA) to induce protease inhibitor gene expression (Koiwa et al. 1997). There are four systemic signals responsible for the translocation of the wound response, which includes systemin, abscisic acid (ABA), hydraulic signals (variation potentials) and electrical signals (Malone and Alarcon, 1995). These signal molecules are translocated from the wound site through the xylem or phloem as a consequence of hydraulic dispersal. The plant systemin an 18-mer peptide has been intensely studied from wounded tomato leaves which strongly induced expression of protease inhibitor (PI) genes. Transgenic plants expressing prosystemin antisense cDNA exhibited a substantial reduction in systemic induction of PI synthesis, and reduced capacity to resist insect attack (McGurl et al. 1994). Systemin regulates the activation of over 20 defensive genes in tomato plants in response to herbivorous and pathogenic attacks. The polypeptide activates a lipid-based signal transduction pathway in which linolenic acid, is released from plant membranes and converted into an oxylipin signaling molecule, jasmonic acid (Ryan, 2000). A wound-inducible systemin cell surface receptor with an M(r) of 160,000 has also been identified and the receptor regulates an intracellular cascade including, depolarization of the plasma membrane and the opening of ion channels thereby increasing the intracellular Ca(2+), which activates a MAP kinase activity and a phospholipase A(2). These rapid changes, play a vital role leading to the intracellular release of linolenic acid from membranes and its subsequent conversion to JA, a potent activator of defense gene transcription (Ryan, 2000). The oligosaccharides, generated from the pathogen-derived pectin degrading enzymes i.e. polygalacturonase (Bergey et al. 1999) and the application of systemin as well as wounding have been shown to increase the jasmonate levels in tomato plants. Application of jasmonate or its methyl ester, methyl jasmonate, strongly induces local and systemic expression of PI genes in many plant species, suggesting that jasmonate has an ubiquitous role in the wound response (Wasternack and Parthier, 1997). Further, analysis of a potato PI-IIK promoter has revealed a G-box sequence (CACGTGG) as jasmonate-responsive element (Koiwa et al. 1997). The model developed for the wound-induced activation of the proteinase inhibitor II (Pin2) gene in potato (Solanum tuberosum) and tomato (Lycopersicon esculentum) establishes the involvement of the plant hormones, abscisic acid and jasmonic acid (JA) as the key components of wound signal transduction pathway (Titarenko et al. 1997). Recently, it has been shown that the defense signaling in suspensions of cultured cells of Lycopersicon peruvianum by peptide systemin, chitosan and by beta-glucan elicitor from Phytophtora megasperma, is inhibited by the polysulfonated naphtyl urea compound suramin, a known inhibitor of cytokine and growth factor receptor interactions in animal cells (Stratmann et al. 2000a). Levels of ABA have been shown to increase in response to wounding, electrical signal, heat treatment or systemin application in parallel with PI induction (Koiwa et al. 1997). Abscisic acid originally thought to be involved in the signaling pathway is now believed to weakly induce the mRNAs of wound response proteins and a concentration even as high as 100 mM induced only low levels of proteinase inhibitor as compared to systemin or jasmonic acid (Birkenmeiner and Ryan, 1998), suggesting the localized role of ABA. However, it is evident that wound induction and pathogen defense pathways overlap considerably. Expression of wound and JA inducible genes can be positively and negatively regulated by ethylene or salicylic acid (SA), both of which are components of the pathogen-induced signaling pathway (Bent, 1996; Delaney et al. 1994). The expression of thionins in Arabidopsis (Epple et al. 1995) and lectin II in Griffonia simplicifolia (Zhu-Salzman et al. 1998) was elicited by JA but suppressed by ethylene, showing their opposite effects on the wound signaling pathway. Plants sometimes specifically forego one type of defense response for another. Salicylic acid (SA) and its methyl ester (Me-SA) are both defense compounds that potently induce systemic acquired resistance of plants against pathogenic microorganisms (Hunt et al. 1996). However, in response to spider mite infestation, lima bean plants release Me-SA which functions as a volatile attractant of the predatory mite Amblyseius potentillae (Dicke et al. 1990). At the same time, SA itself negatively regulates the OD pathway through inhibition of SA biosynthesis and activity (Korth and Dixon, 1997), indicating that SA may suppress the plant defense response through attenuation of the OD pathway, but its methyl ester positively affects plant defense through another defense mechanism involving tritrophic plant herbivore interaction (Moura and Ryan, 2001). Different jasmonic acid-dependent and independent wound signal transduction pathways have been identified recently and partially characterized. Components of these signalling pathways are mostly similar to those implicated in other signalling cascades which include reversible protein phosphorylation steps, calcium/calmodulin-regulated events, and production of active oxygen species (León et al. 2001). Stintzi et al. (2001) using biochemical genetic approach demonstrated that cyclopentenone precursor of JA, 12-oxo-phytodienoic acid (OPDA), as a physiological signal eliciting defensive response and resistance in the absence of JA. Studies on the effect of UV radiation on early signaling events in the response of young tomato plants to ultraviolet-C (< 280 nm) and UVB/UVA (280-390 nm) radiation induces a 48 kDa myelin basic protein kinase activity in leaves (Stratmann et al. 2000b). In the case of barley plants, ethylene increased the activity of both cell wall bound peroxidases types (ionically and covalently bound), comparable with infestation, which suggests that ethylene is involved in the oxidative responses of (Argandoña et al. 2001) Studies on the induction of PI proteins have indicated a de novo synthesis of proteins such as a Boman Birk protease inhibitor (OsBBPI) from rice which was found to be rapidly induced in seedling leaf in response to cut, exogenous jasmonic acid (JA), and two potent protein phosphatase 2A (PP2A) inhibitors, in a light/dark, time and dose dependent manner but was completely inhibited by cycloheximide (Rakwal et al. 2001). Structure of protease inhibitor genes The gene size and coding regions of the inhibitors are generally small with no introns (Boulter, 1993) and many of these inhibitors are products of multigene families (Ryan, 1990). Bowman-Birk type double-headed protease inhibitors are assumed to have arisen by duplication of an ancestral single headed inhibitor gene and subsequently diverged into different classes i.e. trypsin/trypsin (T/T), trypsin/chymotrypsin (T/C) and trypsin/elastase (T/E) inhibitors (Odani et al. 1983). The mature proteins comprise a readily identifiable ‘core’ region, covering the invariant cysteine residues and active center serines, which are bound by highly variable amino and carboxy-terminal regions. There is a core region of 62 amino acids, both between and within the different classes of inhibitor, within cowpea and with other leguminosae, including azuki bean (Ishikawa et al. 1985), lima bean (Stevens et al. 1974), mung bean (Zhang et al. 1982) and soybean (Odani and Ikenaka, 1976). The average number of amino acid replacements in this region from all pair-wise comparisons show that the differences between the different classes of inhibitor within a species (around 16.5/62 residues) are much greater than the differences within a class between different species (around 11/62 residues). Considering that 18 of the residues in this region are obligatorily invariant for proteins to be classified as Bowman-Birk type inhibitors, these are very high rates of amino acid substitutions. This highlights the problems likely to be encountered in attempting to draw conclusions about the evolutionary history of the rapidly diverging, multigenic protein families from sequences which may be paralogous rather than orthologous. Corrected divergence between pair-wise combinations of sequences calculated according to the method of Perler et al. 1980 revealed that the average divergence between trypsin-specific and chymotrypsin-specific second domains (about 36%) is very similar to that between the first and second domains (about 40%). On an “evolutionary clock” model this would imply that the gene duplication leading to T/T and T/C families occurred very close to the duplication, leading to the appearance of the double-headed inhibitors and that the number of silent substitutions has reached saturation in all these genes (Hilder et al. 1989). Analysis of the winged bean Kunitz chymotrypsin inhibitor (WCI) protein shows that it is encoded by a multigene family that includes four putative inhibitor coding genes and three pseudogenes. The structural analysis of the WCI genes indicates that an insertion at a 5' proximal site occurred after duplication of the ancestral WCI gene and that several gene conversion events subsequently contributed to the evolution of this gene family (Habu et al. 1997). The 5' region of the pseudogene, WCI-P1 contains frameshift mutations, an indication that the 5' region of the WCI-P1 gene may have spontaneously acquired new regulatory sequences during evolution. Since, gene conversion is a relatively frequent event and the homology between the WCI-P1 and the other inhibitor genes WCI-3a/b is disrupted at a 5' proximal site by remnants of an inserted sequence, the WCI-P1 gene appears to be a possible intermediate that could be converted into a new functional gene with a distinct pattern of expression by a single gene-conversion event (Habu et al. 1997). Molecular evolution of wip-1 genes from four Zea species show significant heterogeneity in the evolutionary rates of the two inhibitory loops, in which one inhibitory loop is highly conserved, whereas the second is diverged rapidly. Because these two inhibitory loops are predicted to have very similar biochemical functions, the significantly different evolutionary histories suggest that these loops have different ecological functions (Tiffin and Gaut, 2001). The 3’ends of the sequences are comprised of alternating purine-rich and pyrimidine-rich traits of nucleotide. The first of these purine-rich trait occurs at the C-terminal region of the coding sequence. Mutations within this region (deletion/additions or base changes) give rise to substitution amongst the A/G rich codons, Asp, Glu, Lys, Asn and termination codons and also certain specific motifs within them appear to be relatively conserved, therefore likely to be of functional significance. This obviously applies to the cononical polyadenylation signals in the third and second purine-rich traits and the 3’region has regulatory elements which are dependent on a higher order of structure than the specific base sequence. X-ray crystallography of Bowman-Birk inhibitors suggests that this termini has no role in the interaction of the inhibitor with its target enzyme (Suzuki et al. 1987). Substitutions and deletion/additions appear to be very feasible in this region, provided that there is a cleavable serine or asparagine residue within 10-20 amino acids of the first cysteine (Ryan, 1990). The inhibitors are synthesized as precursors from which the leader sequence is cleaved and a long trait of leader-encoding sequence is present in soybean genomic clones (Hilder et al. 1989). There is no significant homology in this region to other seed-expressed protein leader sequences, other than a high representation of hydrophobic residues. Multiple potential initiator codons are a common feature of legume seed protein genes exemplifying the high degree of evolutionary novelty which appears to be tolerated within such seed specific secondary compound genes (Hilder et al. 1989). Analysis of oryzacystatin OC-I has revealed the presence of two introns; the first a 1.4 kbp region between Ala 38 and Asn 39 and a second region of 372 bp in the 3’ non coding region (Kishimoto et al. 1994) and a second oryzacystatin, OC-II present on chromosome 5 also has introns in the same positions (Kondo et al. 1991) thus suggesting deviation from the earlier PIs which lacked introns. Developing insect resistant transgenic plants expressing Pls A large number of protease inhibitor genes with distinct modes of action have been isolated from a wide range of crop species. Development of transgenic crops have come a long way from the first transgenic developed by Hilder et al. 1987. Considering the high complexity of protease/inhibitor interactions in host pest systems and the diversity of proteolytic enzymes used by pests and pathogens to hydrolyze dietary proteins or to cleave peptide bonds in more specific processes (Graham et al. 1997), the choice of an appropriate proteinase inhibitor (PI) or set of PIs represents a primary determinant in the success or failure of any pest control strategy relying on protease inhibition. Firstly, the choice of suitable PIs should be based on a detailed understanding of the biological system assessed. Based on our current knowledge about the use of specific inhibitors in the study and control of various metabolic pathways, many PIs have been used to create transgenic crop plants as shown in Table 1 and many more inhibitors are also being isolated with divergent modes of action against different pest species (Table 2). Resistant biotypes of insects may evolve after prolonged exposure to selection pressure that is mediated by an insecticidal protein or plant resistance gene (Sparber, 1985). Unless the biotechnology strategy is designed and implemented to overcome these problems, it will become ineffective in due course like any pesticide based management strategy. Second point to consider would be the targeted expession of PIs in response to pest attack. This will be controlled by using inducible promoters, such as those of PI-II85 and TobRB7, that are activated at the site of invasion by pests, pathogen and nematodes, respectively (Opperman et al.1994). An ideal promoter should be highly responsive to invasion of the host plant by a pest, or regulated by inducers just prior to pest attack. The promoter should be sufficiently active to mediate a substantial defense, specially localized to the site of pest invasion. Suitable promoters such as those regulated in response to pest invasion can be identified using promoter trapping techniques (Babiychuk et al. 1997). Despite these promising developments, the general usefulness of recombinant PIs in plant protection still remains to be demonstrated. The inhibitory spectrum of PIs is usually limited to proteases in one of several mechanistic classes, leaving free proteases in the surrounding medium after inhibition (Barrett, 1994). Due to a progressive adaptation of plant pests to the continuous occurrence of PIs in the diet, the inhibitory spectrum of protein inhibitors against the extracellular proteases of several pests is even more limited, being often restricted to the family level (Michaud et al. 1995b; Visal et al. 1998). Non-target proteases, that may allow metabolic compensation of inhibited proteolytic functions, (Jongsma and Bolter, 1997) may also challenge the structural integrity of several PIs and thus potentially affect their effectiveness in vivo (Michaud, 1997). Recently it has been shown that the presence of large amounts of inhibitors including soybean Kunitz inhibitor (Bown et al.1997) in the diets of economical pests has made insects to adapt and produce proteases which are insensitive to the action of host plant inhibitors and the ingested PIs activate these genes (Dennis et al. 1994). As a result, pest control using PIs in transgenic plants requires the isolation of inhibitors that are active towards these insensitive proteases (Jongsma et al.1996). One can search for active inhibitors among naturally-occurring peptides (Gruden et al. 1998) or can engineer inhibitors in such a way that they will acquire activity against the "PI insensitive" protease. Engineering of inhibitors can be performed in two distinct ways: 1) based on structures of the inhibitor-protease complex, predictions can be made on mutations that will enhance binding (Urwin et al. 1995), but lack of data on these complexes for insect proteases makes this procedure rather tough. However, it may be more appropriate to simply generate large arrays of mutants in the region of the inhibitor protein contacting the protease. The powerful method of phage display can subsequently be used to select the strongly binding mutants. Large number of PIs have been subjected to phage display as a result of which inhibition constant (Ki) for target protease which was initially poor (millimolar to micromolar) have been greatly improved. Some of the examples belonging to serine class are serpin (Pannekoek et al. 1993) and kazal from human (Rottgen and Collins, 1995), E.coli ecotin (Wang et al. 1995), proteinase inhibitor II (PI-II) from potato (Jongsma et al. 1995), chicken cystanin (Tanaka et al. 1995) and soybean phytocystanin (Koiwa et al. 1998) belonging to cysteine class. The improvement of plant PIs by phage display is still an infant stage to be commercially important. Insect midgut contains an estimated 1020 different proteases (Bown et al. 1997) which are differentially regulated and all cannot be inhibited by plant’s PIs (Broadway, 1997). Therefore, to achieve an effective pest control strategy it is very important to achieve different inhibitors expression in a concerted manner. Concluding Remarks The availability of diverse genes from different plant species makes it a possibility to use one or more genes in combination, whose products are targeted at different biochemical and physiological processes within the insect. These packages will not only contain protease inhibitor genes but also lectins, alpha-amylase inhibitors, or other plant genes encoding insecticidal proteins. This technology may not replace the use of chemical pesticides in near future but effectively complement it. The use of recombinant PIs may also be an attractive way to protect plants from fungal, bacterial and viral pathogens. Currently, two principal strategies are proposed to engineer effective pest control in plants: ectopic expression of pesticidal proteins, and induction of the plant natural defensive response. At present, screening gene pools without taxonomic constraint can help identify novel insecticidal determinants, but in future this approach will be augmented by directed in vitro molecular evolution (Koiwa et al. 1998). Given the number of pesticidal proteins that are involved in host plant defense, it is presumed that effective pest control by this strategy will result from the co-expression of numerous determinants, each of which could be custom engineered by directed molecular evolution to maximize its effectiveness against specific pests. However, in future non-scientific issues such as regulatory approval, propriety rights and public perception will be decisive in releasing crop plants produced by genetic engineering using recombinant DNA technology. References
Note: EJB Electronic Journal of Biotechnology is not responsible if on-line references cited on manuscripts are not available any more after the date of publication. Supported by UNESCO / MIRCEN network © 2002 by Universidad Católica de Valparaíso -- Chile The following images related to this document are available:Photo images[ej02015t1.jpg] [ej02015t2.jpg] |
| |||||||||

{kind=link}
{kind=link}