 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Electronic Journal of Biotechnology, Vol. 5, No. 2, August, 2002 Transfer and expression of the genes of Bacillus branched chain alpha-oxo acid decarboxylase in Lycopersicun esculentum Gefu Wang-Pruski*1 and Aladar A. Szalay2 1Department of Plant and
Animal Sciences, Nova Scotia Agricultural College, PO Box 550, Truro, Nova Scotia,
B2N 5E3, Canada Tel: 902 893 6247 Fax: 902 897 9762 E-mail: gwangpruski@nsac.ns.ca Financial support: Grants from Canadian Bacterial Diseases Network, Natural Science and Engineering Research Council of Canada, and American Gene Therapy Inc. Received October 10, 2001 Code Number: ej02025 Abstract Engineering of higher plants for increased cold tolerance requires a chemical modification of membrane fluidity in both organelles and cytoplasm of plant cells. A small number of microorganisms use branched chain fatty acids as their membrane constituents to maintain membrane fluidity, instead of unsaturated fatty acids. One of the key enzymes facilitating synthesis of branched chain fatty acids in Bacillus subtilis 168s is the branched chain alpha-oxo acid decarboxylase. To examine the role of branched chain fatty acids in plants and the potential for low temperature tolerance, the A and B genes encoding the alpha and beta polypeptides, respectively, of the branched chain alpha-oxo acid decarboxylase were introduced into the genome of tomato plants. The mas promoters in the plant expression vector system facilitated the expression of the A and B genes. Transgenic plants were regenerated and shown to produce both the alpha and beta polypeptides. Comparative analysis of a small number of transgenic tomato plants and non-transformed control plants grown at 4ºC showed enhanced cold tolerance in the transformed plants. These findings, if confirmed by a larger scale analysis, suggest a potential role for branched chain fatty acids as a protective mechanism for growth of plants under sub optimal temperatures. Keywords: branched chain alpha-oxo acid decarboxylase, cold tolerance, gene expression, transgenic tomato plants. Article Physiological manifestations of environmental stress in plants often implicate cell membranes as the primary site of injury (Orvar et al. 2000). At low temperatures, a decrease in membrane fluidity leads to a decrease in the activity of membrane-bound enzymes and loss of semi permeable membrane properties (McMurchie and Raison, 1979; Levitt, 1980). This is due to the transition of membrane lipids from a fluid liquid-crystalline phase to a viscous gel crystalline phase (Cropp et al. 2000). The acyl chain region, comprising the hydrophobic domain of most membrane lipids, determines membrane fluidity. This domain contains fatty acids, either straight chain or branched chain families. The branched chain fatty acid family contains iso-, anteiso- and omega-alicyclic fatty acids with or without a substitution (unsaturation and hydroxylation) (Kromovar et al. 1994; van der Hoeven and Steffens, 2000; Graham and Eastmond, 2002). Branched chain fatty acids in cell membranes contribute to the control of membrane fluidity (Kromovar et al. 1994). In the genus Bacillus, the majority of fatty acids contributing to membrane fluidity are branched chain fatty acids (Jackowski et al. 1991). Iso-acyl phosphatidylcholine, one group of branched fatty acids, has a melting point (Tm) ranging from 18-28ºC below that of the corresponding normal saturated acyl phosphatidylcholine (Silvius, 1982). Studies of cold tolerant microorganisms suggest that the fatty acid composition of their cell membranes differs from that of their counterparts adapted to warmer ecological niches. Membrane fatty acid analysis of Antarctic bacteria showed that branched chain fatty acids were predominant in the membrane fatty acids in the psychrophiles (Rotert et al. 1993). Deep-sea barotolerant bacteria contain branched chain fatty acids as a major component of membrane lipids (Kaminura et al. 1993; Nichols et al. 2000). Legionella shakespearei sp. isolated from cooling tower water contained branched chain fatty acids as the major membrane lipid component (Verma et al. 1992). Other reports show that some Listeria and Legionella cells are resistant to cold temperature due to the increased branched chain fatty acid concentration (Benson et al. 1996; Juneja et al. 1998). The proportion of anteiso- and iso- branched chain acids is increased by reduced temperature, rather than induction of fatty acid unsaturation. These findings suggest that membrane fluidity may be enhanced at low temperatures by increasing the ratio of branched chain fatty acids in cell membranes. The biosynthetic pathway of straight and branched chain fatty acid catabolism in higher plants has been recently reviewed (Graham and Eastmond, 2002). This review pointed out the advances in the understanding of branched chain amino acid catabolism have provided convincing evidence that mitochondria play an important role in this process. The biosynthetic pathway of branched chain fatty acids in bacteria is better understood (Kaneda, 1991). Only some bacteria, a few protozoa, and a few fungi use branched chain fatty acids as their membrane components (Kaneda, 1977; Kaneda, 1991). These organisms use branched chain alpha-oxo acids related to valine, isoleucine and leucine as precursors. These alpha-oxo acid substrates are decarboxylated to yield the precursor compounds for fatty acid synthesis reactions. The decarboxylase, named as branched chain alpha-oxo acid decarboxylase (BCOADc), has been found to be essential in fatty acid synthesis from alpha-oxo acids (Oku and Kaneda, 1988). BCOADc is a heterodimer consisting of two alpha subunits and two beta subunits, with a molecular weight of 144.4 KDa. Previously, three genes encoding branched chain alpha-oxo acid dehydrogenase complex (EC1.2.1.25) were cloned and sequenced from B. subtilis 168s (Wang et al. 1993). The first two genes encode alpha subunit (36310 Da) and beta subunits (35893 Da) of the BCOADc; the third gene encodes the dihydrolipoyl acyltransferase and is considered to be a part of the dehydrogenase complex (Wang et al. 1993). The BCOADc essential in branched chain fatty acid synthesis from alpha-oxo acid may be a free form of the dehydorgenase of the branched chain alpha-oxo acid dehydrogenase complex (Wang et al. 1993). In this report, we present the transfer and expression of the first two genes that encode branched chain alpha-oxo decarboxylase (BCOADc) in the tomato plant, L. esculentum, and the potential for enhanced biosynthesis of branched chain fatty acids in plants to increase cold tolerance capability. The mannopine synthase (mas) promoter is a dual promoter containing two promoter (P1, P2) activities in opposite directions (Velten et al. 1984; Langridge et al. 1989). In this study, two genes, the A gene, encoding the alpha subunit, and the B gene, encoding the beta subunit of the decarboxylase, were placed under the P1 and P2 promoters, which permit the induction and expression of both genes in the same plant cell. Using the Agrobacteriun mediated T-DNA transfer method, we generated tomato plants which carried single or multiple copies of the foreign genes. Transformed plants were shown to produce the correct transcripts, and the alpha and beta polypeptides of the BCOADc enzyme. Comparative analysis of a smaller number of transgenic tomato plants grown at 4oC showed these plants possessed enhanced cold tolerance compared to non-transformed control plants. These initial studies suggest that the introduction of genes essential for branched chain fatty acids biosynthesis into plants may provide a new approach in generating low temperature tolerant plants. Materials and Methods Experimental materials. Plasmid pPCV701
containing mannopine synthase gene promoters, the dual mas promoters
(Koncz et al. 1987), was used to drive the expression of the
BCOADc A and B genes in tomato plants. The new construct was named as pPCV701-AB
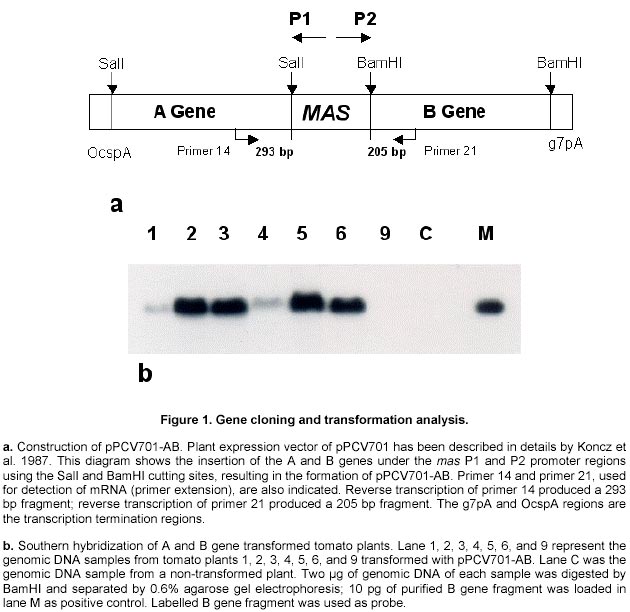
(Figure 1a). Plasmid pBluescript
II KS +/- was used as a cloning vector for cloning of the PCR products and for
over expression of the alpha and beta polypeptides of BCOADc in E. coli
DLT101. E. coli DLT101 (LE392 Cloning primers. The primer sequences for gene cloning and primer extension experiments are listed as following. A1:
5'-CGGTCGACATGAGTACAAACCGACATCA-3' B1:
5'-CGGGATCCATGTCAGTAATGTCATATAT-3' ASD:
5'-CGGTCGACAAGGAGGGCTTGAATGAGTA-3' Primer
14: 5'-TACGGCAATACATAATCC-3’ The A and B genes encoding the BCOADc enzyme were amplified by PCR using primer A1 and A2, B1 and B2, respectively, from the genomic DNA of B. subtilis 168s. Primer A1 and A2 were designed to include a SalI restriction cutting site. Primer B1 and B2 were designed to include a BamHI cutting site. For over expression of the alpha and beta polypeptides in E. coli DLT101, the A and B genes were amplified by PCR using primer ASD and A2, and primer BSD and B2, respectively. Both the ASD and BSD primers contained Shine-Dalgarno (SD) sequences. The Primer 14, designed based on the DNA sequence in the A gene, was used for the detection of the mRNA transcripts of the A gene; Primer 21, the sequence from the B gene, was used for the detection of the mRNA transcripts of the B gene. Transformation
of tomato plant var. Roma. Agrobacterium mediated cotyledon
transformation was used for introducing genes into tomato plants. Surface sterilized
seeds of tomato L. esculentum were germinated on 1/2 MSO medium (2.3 g
Gibco M8 salts, 3% sucrose, 1 H B5 vitamins). Before appearance of true leaves,
the cotyledons were collected, cross sectioned and immediately floated in 5 ml
of MSO liquid medium with 5 µl cultured Agrobacterium (OD = 1.0) and 375
µM acetosyringone. After incubation for 5 min, they were placed upside down on
D1 medium [1 x MS salts (Sigma), 3%
glucose, 1 H B5 vitamins, 1 mg/L zeatin, 0.22% gelrite, pH 5.8]. Two days later,
these cotyledons were transferred to D1 medium containing 100 mg/L kanamycin and
300 mg/L claforan. After three to four weeks, shoots with true meristems were
cut from the cotyledon tissues and transferred to rooting medium (1 x MS, 2% glucose,
0.22% gelrite). Well-rooted plantlets were then transferred into wet soilless
media (Metro-Mix #290), and shaded for five days before moving to the greenhouse.
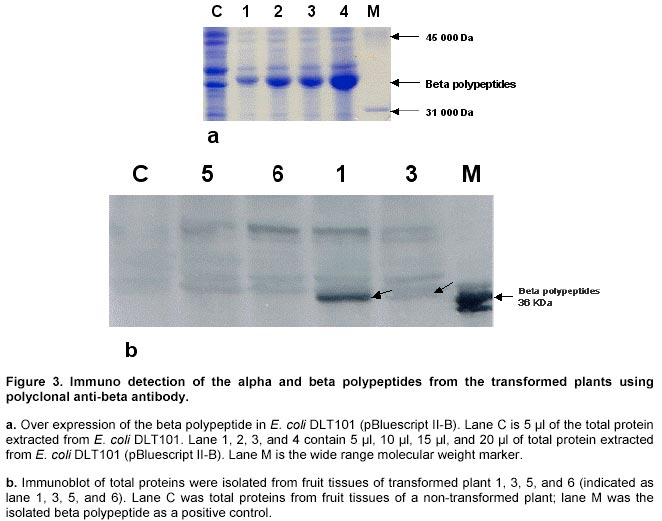
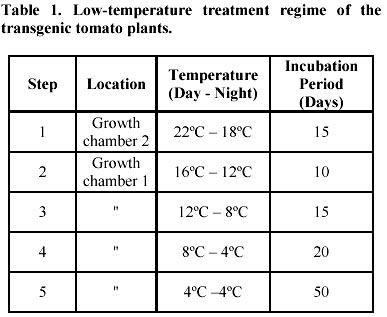
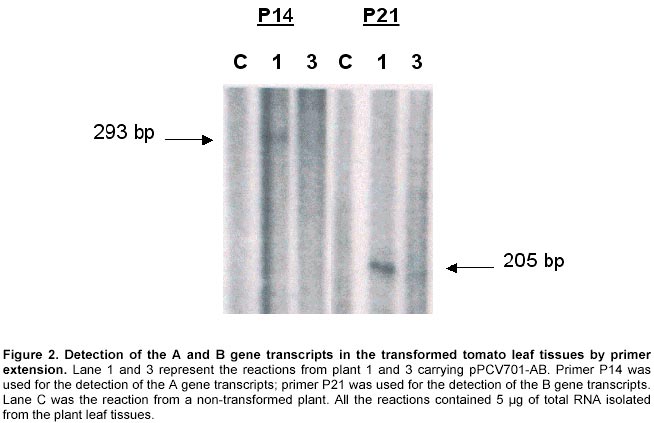
Detection of gene integration and gene expression. Plant genomic DNA was isolated from leaf tissues (Doyle and Doyle, 1990), digested with selected restriction enzymes and separated on 0.6% agarose gels. Southern hybridisation was followed by transferring the DNA to nylon membranes (MagnaGraph, MSI) and hybridised with alpha-32P-dCTP labelled B gene fragments as probes. Detection of mRNA transcripts was achieved by primer extension. The primer extension method was used to map and quantitate the 5' termini of RNA and to detect mRNA. The test RNA was hybridised with an excess of an end-labelled single-stranded DNA primer. Reverse transcriptase was then extended the primer to produce cDNA complementary to the RNA template. The length of the resulting end-labelled cDNA, as measured by electrophoresis through a polyacrylamide gel under denaturing conditions, reflected the distance between the end-labelled nucleotide of the primers and the 5' terminus of the RNA. The yield of cDNA was proportional to the concentration of the target mRNA sequences in the total preparation. In this study, oligonucleotide primers 14 and 21 were end-labelled by gamma-32P-ATP using T4 polynucleotide kinase. The quality of the labelled primers was checked using gel electrophoresis (15% minipolyacrylamide gel). To initiate the reverse transcription reaction, 5 µg of total RNA, isolated from leaf tissues of the plants, grown in greenhouse for four weeks after transplanting, was dissolved in 3 µl of water and mixed with 1 µl (5 pmoles) labelled primer. The mixture was denatured in a 90ºC water bath for 1 min, then quickly transferred to a 42ºC heat block. Buffer I, 4 µl, containing 140 mM beta-mercaptoethanol, 100 mM Tris-HCl, pH 8.3, and 0.7 M KCl, was added to the tube and the mixture incubated at 42ºC for an additional 15 min. When this incubation was completed, the extension mix (12 µl of Buffer II with 10-20 units of AMV reverse transcriptase) was added and incubated for 30-40 min at 42ºC. Buffer II contains 133 mM Tris-HCl, pH 8.3, 16 mM MgCl2, and 1.3 mM dNTPs. The reaction was stopped by adding 1/10 volume of DNA loading buffer, and the DNA samples were denatured in boiling water, and 1/3 of the reaction was loaded on a 5% polyacrylamide gel. The gel was dried and placed under X-ray film for exposure at –70ºC for 1-2 days. Production of anti-beta peptide antibody. The A and B genes were amplified by PCR from genomic DNA of B. subtilis 168s using primer ASD and A2 for the A gene, primer BSD and B2 for the B gene, respectively. Amplified gene fragments were ligated into E. coli expression vector pBluescript II KS +/-, and resulted in pBluescript II-A and pBluescript II-B. Both the pBluescript II-A and pBluescript II-B plasmid constructs were transformed into E. coli DLT101. IPTG 0.2 mg/ml was added into the E. coli DLT101 culture (100 ml, OD600 = 1.0) and incubated for an additional 1 hr. The induced cells were collected and lysed in protein sample buffer (50 mM Tris-HCl, pH 6.8; 10% glycerol, 2% SDS, 2mg/ml bromophenol blue, 1% beta-mercaptoethanol). The supernatant was boiled and loaded onto a 7.5-25% gradient SDS-polyacrylamide gel and separated at 35 mA for 16 hrs in Tris-glycine buffer. Figure 3a demonstrates the induction of the beta polypeptide in E. coli DLT101 cells. After completion of the electrophoresis, gels were stained in 0.2% Coomassie brilliant blue for 10 min. Protein bands were identified and the band of interest was cut out from the gel, crushed into small pieces and loaded into a glass tube (8 x 100 mm). Dialysis tubing was fixed at the bottom end of the glass tube and proteins were eluted from the gel by electrophoresis. The purified protein was confirmed by SDS-PAGE and was directly used for the preparation of antibodies in rabbits. New Zealand white females, 3 months old and approximately 3-4 kg in weight, were used for the preparation of antibodies. The immunization process was divided into four steps: antigen injection, first boost, second boost, and the third boost. Sera were collected 15 days after each boost. To collect the sera, blood samples were centrifuged at 3,000 rpm for 30 min, and the clear serum was harvested. Twenty-five ml of saturated ammonium sulphate solution (pH 7.8 by 2 N NaOH) was added (drop wise) to 50 ml of serum sample with constant stirring. The mixture was stirred for an additional 2 to 3 hrs at room temperature then centrifuged for 30 min at 1,400 g. The resulting pellet was dissolved in 5 ml of PBS buffer. Ammonium sulphate from the precipitate was removed by dialysis against PBS buffer for 2 days at 4ºC. The quality of antibodies was checked by immunoblotting with a series of diluted protein antigens. Immunoblot analysis. Leaf or fruit tissue from transgenic tomato plants were sterilized and induced on MS + 2,4-D (0.5 mg/L) medium for maximum induction of mas dual promoter activity. Tissues were collected after 3 days induction, homogenized in extraction buffer (50 mM Tris-HCl, pH 7.5, 2 mM EDTA, 0.5 mM EGTA, 1mM PMSF, 1% Triton X-100) and then total protein suspension was collected by centrifugation at 2,000 g. The suspensions were mixed with 1/4 volume of sample buffer (50 mM Tris-HCl, pH 6.8, 10% glycerol, 2% SDS, 2mg/ml bromophenol blue, 1% beta-mercaptoethanol). The samples were boiled for 3 min and 100 µl aliquots of each were loaded on a 3 mm 7.5-20% gradient polyacrylamide gel. The protein gels were electro-transferred from polyacrylamide gels to PVDF membranes at 200 mA for 3 hrs. Before immunoblotting was started, PVDF membrane containing protein samples was immersed in 100% methanol for 5 min, rinsed with water, then incubated in TBST buffer (10 mM Tris-HCl, 150 mM NaCl, 0.05% Tween 20, pH 8.0) for 5 min. The immunoblotting process was initiated by incubation of the membrane in freshly made blocking solution containing TBST + 1% BSA + 20% Fatal Calf Serum (Sigma) for 1 hr with shaking. The blocking solution was then replaced with TBST containing the antibody against the target protein at a dilution of 1:2,000 to 1:10,000 and incubated at room temperature for 1 hr with shaking. After incubation, the membrane was washed 3 times and the second antibody (goat anti-rabbit alkaline phosphatase conjugate, Sigma) was added in TBST with a dilution of 1:5,000, and incubated for 30 min as before. The membrane was washed as before and treated for 2 min in alkaline phosphate buffer. Membrane in BCIP/NBT solution (Sigma) was incubated in the dark for 2 to 60 min and the colour reaction was terminated by removal of colour reagents and rinsing the membrane in water. Cold tolerance test of the transformed plants. Seeds of transformed plant 1 were generated on ½ MSO (2.3 g Gibco M8 salt, 3% sucrose, 1 x B5 vitamins, 0.22% gelrite, pH 5.8) in the presence of 100 mg/L kanamycin. Seedlings with undeveloped roots were discarded. The generated seedlings were confirmed by Southern hybridisation and grown to six inches in height in a growth chamber (Table 1). Control plants were generated from seeds at the same time. Plants for low temperature testing were arranged in three replicated groups. Each group contained three plants, one control non-transformed plant and two transformed plants that contained the A and B genes. All plants were placed in a growth chamber for 15 days under optimal growth conditions, and then transferred to another growth chamber where the low-temperature regime was initiated (Table 1). Results and Discussion Construction of plant expression vectors encoding BCOADc genes Primers A1 and A2, containing a SalI cleavage site were used to excise the A gene from the genomic DNA using PCR. Primers B1 and B2, containing the BamHI cleavage sites were used to excise the B gene from the genomic DNA in a similar manner. The two PCR products (A and B) were independently subcloned into pBluescript II KS +/- , and the genes were sequenced to confirm the sequence accuracy. Further, the A and the B genes were subcloned into the plant expression vector pPCV701 under the control of the dual mas promoter, P1 and P2, respectively. The resulting vector, pPCV701-AB, is shown in Figure 1a. Southern hybridisation analysis Genomic DNA from pPCV701-AB transformed plants 1, 2, 3, 4, 5, 6 and 9 was isolated and 2 µg of the genomic DNA was digested with BamHI restriction enzyme. The digested DNA samples were separated by agarose gel electrophoresis and transferred to nylon membrane. Southern hybridisation was performed with the B gene fragment labelled with alpha 32P-dCTP as the probe (Figure 1b). As shown in Figure 1b, positive signals were detected from plants 1, 2, 3, 4, 5, and 6. Plant 9, grown under kanamycin selective conditions, did not carry the transformed gene construct. Band intensity reflects different copy numbers of the integrated foreign genes in the transformed plants. Copy number of each transgenic plant was analysed using HindIII restriction enzyme, the single cutting site in the A genes. Southern hybridisation results indicated that single and multiple insertions occurred in these transgenic plants. Plants 1 and 4 produced single gene inserts while plants 2, 3, 5, and 6 contained multiple copy gene inserts (data not shown). The multiple copies of the gene were inserted either in one location as arrays, or inserted in multiple locations. Transcriptional analysis of the A and B genes of BCOADc Primer extension was selected for the detection of the A or B gene transcripts in individually transformed plants (Figure 2). Primer P14 was designed for the A gene which should produce a reverse transcribed product of 293 bp; primer P21 was designed for the B genes which should produce a 205 bp reverse transcribed product. Using equal amounts of total RNA isolated from leaf tissue, reactions using primer 14 and primer 21 produced two reverse transcribed cDNA fragments. Sizes of 293 nucleotides and 205 nucleotides, calculated based on the DNA ladder, were found and correspond to the expected sizes (Figure 2). The strongest cDNA signal for both the A and B gene transcripts were detected in plant 1. Weaker signals were found in plant 3 for both reactions. No visible signal were observed in the other plants (data not shown). Previous Southern hybridisation data has shown that plant 1 carried a single copy of the A and B gene construct; plant 3, however, carried multiple copies of the transgenes. The fact that the plant 1 produced more transcripts than plant 3 indicated that higher copy number of transgenes might not correspond to higher transcriptional levels. Other factors, such as the location of the integrated transgene in the chromosome may play an important role in gene activation. This observation is supported in early studies by Hobbs et al. 1993 and Grevelding et al. 1993 that transgene copy number can positively or negatively affect transgene expression. Production of the polypeptides in E. coli and their antibodies in animals Using the ASD and A2, and BSD and B2 primer, the A and the B genes were amplified by PCR. The amplified PCR products acquired the SD sequence 5-9 bp upstream of the ATG start codons. They were then subcloned to the T7 promoter of the plasmid pBluescript II KS +/-, resulting in the formation of pBluescript II-A and pBluescript II-B. Both plasmids were then transformed into E.coli DLT101, and over production of the alpha and beta polypeptide subunits were performed. Successful induction of the beta polypeptide is shown in Figure 3a. The protein band was excised from the polyacrylamide gel and the polyclonal anti-beta antibody was successfully produced. Quantitative and qualitative assays of the generated antibodies were performed by immunoblotting the beta protein with a series of dilutions of the antibody. Specific bands were obtained from 200-, 500-, 1,000- and 2,000-fold diluted polyclonal antibody mixtures. The 2,000-fold dilution, which displayed the least background and the clearest pattern of protein, was selected for Western (immunoblot) analysis of transgenic plant proteins (data not shown). The same procedure was carried out for the alpha polypeptide. However, the level of the alpha protein induction was much lower than the beta polypeptide and the quality of the anti-alpha antibody was not specific enough for immunoblot analysis. Therefore, the immunoblot detection of the transformed tomato plants was performed using anti-beta antibody (Figure 3). Immuno blot analysis of BCOADc gene products in tomato tissues Since the mas promoter can be induced by the plant growth hormone auxin (Langridge et al. 1989), green fruit tissues were sliced and incubated prior to protein extraction to maximize the promoter activation. Four independent pPCV701-AB transformed plants (1, 3, 5, 6) were examined (Figure 3b). Using the anti-beta antibody, plant 1 showed a strong blotting signal at the position identical to the beta polypeptide, indicating the presence of the beta polypeptide in the tissue. Plant 3 showed a weaker signal at the same position. There was no obvious signal being detected from plants 5 and 6. However, immunoblot analysis detected one band at higher molecular weight (Figure 3b) from all four transformed tomato plants, but no such signal was present for the control non-transformed plant (Figure 3b). Molecular weight examination showed that the band was around 72 KDa. Since the alpha and beta subunits of the BCOADc have a molecular weight of 36310 Da and 35893 Da, respectively (Wang et al. 1993), we believe the signal of 72 KDa consists of the hetero dimer of one alpha and one beta polypeptide subunit. Since the samples were separated on SDS-PAGE gel, it is difficult to explain the formation of the dimmer subunit. It is difficult to predict that if the band at 36 KDa contained only the beta polypeptide or both the two alpha and the beta polypeptides, since their molecular weight is very close. Similar experiments were carried out to detect the alpha and beta subunits in leaf tissues of these transformed tomato plants. The same signal was shown in plant 1. However, no signal was detected from any other plants. Using the mas promoter, Koncz et al. 1987 reported that alpha and beta subunits of the bacterial luciferase enzyme were produced in a similar amount in tobacco plants. Both subunits could also be properly assembled in plant cells (Koncz et al. 1987). The assembly of the alpha and beta subunits of BCOADc in tomato plants in this study needs to be further confirmed. In plants, fatty acid beta-oxidation takes place mainly in peroxysomes, and the cellular location of the enzymes involved in the catabolism of branched chain amino acids has not been definitely assigned. A recent study suggests that the enzymes are located in glyoxysomes, but no immunoreactive bands were detected in mitochondrial fractions (De Bellis et al. 2000). Growth of transgenic plants under low temperature conditions Seeds from plant 1 were germinated on ½ MSO medium containing 100mg/L kanamycin. Of 20 seeds germinated, four did not develop root system. Southern hybridisation experiments were used to confirm that the test plants carried the transgenes. Test plants were grouped into three groups. Each group contained one control, non-transformed plant, and two transformed plants. The seedlings were first transferred into pots and grown under optimal condition (22ºC-18ºC, day-night) in growth chamber 2 (Table 1, Step 1). After 15 days, the plants were fully developed as shown in Figure 4a. They were then transferred to growth chamber 1 where the low temperature treatment took place (Table 1). Temperature treatment was performed by gradual temperature decrease. The temperature was decreased at 4ºC intervals, starting at 16/12ºC, day/night (Step 2). There was always 4ºC day/night temperature difference until the temperature reached 4ºC (Table 1, Step 3, 4, 5). The incubation period (Days) at each temperature setting was determined based on if morphological changes had occurred (Table 1). When the plants were first transferred to 16/12ºC (day/night) for ten days (Table 1, Step 2), all the plants showed normal growth. When the temperature was lowered to 12/8ºC (Step 3), and then further to 8/4ºC (Step 4), stems started to turn dark purple and leaves became dark green with dark purple spots. Five days after the temperature was lowered to 4/4ºC (Step 5), wilt symptoms began to appear on the control plants (Figure 4b-1). All three control plants were dead after 10 days exposure to 4ºC, whereas the transformed plants appeared to be alive. After twenty days of growth under these conditions, one of the transformed plants wilted. A second transformed plant showed similar damage 35 days after cold treatment. The remaining transformed plants were able to tolerate 4ºC for 50 days and did not wilt. Figure 4 shows the test plants from one of the three groups. Figure 4a shows the plants on day 15 and Figure 4b on day 23. In this group, the two transformed plants survived the 4ºC treatment for 50 days, while the non-transformed plant was dead after 10 days. During the whole testing period, plants were immediately moved to another growth chamber (with optimal conditions) when wilt symptoms started to appear. None of the control plants recovered. The two wilted transformed plants showed recovery after two weeks under optimal growth conditions. All other four transformed plants that survived at 4ºC for 50 days were moved back to chamber 2 after the treatment was completed (Step 5). Plants all produced new shoots and leaves. Most of the younger leaves survived the cold treatment and recovered while older leaves were damaged and did not recover. When plant cells are exposed to temperature near 0ºC, viability declines. Overall photosynthesis, photosynthetic electro transport, and phosphorylation activities are all diminished (Tomczak et al. 2002). Also, potassium ions, glutamate, and pteridines are released from cells. These low-temperature-dependent phenomena are irreversible, and even when the cells are placed under temperatures suitable for growth, only partial recovery or none at all were observed (Fernandez Murga et al. 2001). Critical temperatures for irreversible damage of cells depend on growth temperature, suggesting that the temperature-dependent alteration of membrane lipid may be involved in the susceptibility to low temperature. A mechanism has been proposed for these low temperature-induced irreversible phenomena. The plasma and thylakoid membranes are barriers for ions and small molecules, whereas the outer membrane of the cell envelop is permeable to them. At optimal growth temperatures, the plasma membrane and thylakoid membranes are both in the liquid crystalline state and are impermeable to ions and small molecules. With a decrease in temperature, the thylakoid membranes go into the phase separation state and become permeable to ions and small molecules. Under these conditions, physiological activities such as photosynthesis and photosynthetic ATP formation are reversibly diminished (Murata and Fork, 1975; Murata and Sato, 1983). With a further decrease in temperature, the plasma membrane enters the phase separation state and becomes permeable. Under these conditions, ions and small molecules in the cytoplasm leak out, and those in the surrounding medium leak in. This diminishes cellular metabolism leading to cell death. When the temperature is raised to that which supports growth in unchilled cells, the concentration of ions and small molecules are not recovered, resulting in irreversible damage of physiological activities of the cell. Thus, a phase transition in the plasma membrane is directly related to low temperature damage. The effect of the alpha and beta polypeptides of BCOADc enzyme was tested by comparative analysis of transformed and non-transformed control tomato plants, subjected to cold temperature regime. Among the plants tested, the three non-transformed plants showed symptoms of damage much earlier than the plants containing the BCOADc A and B genes. The preliminary findings of this project would be strengthened by additional experiments. Detailed analysis of cell membrane fatty acid composition could be carried out to demonstrate the synthesis and the incorporation of branched chain fatty acids into cell membranes. We initiated the analysis of the fatty acid composition from tomato leaf and fruit tissues using gas chromatography, however we did not find significant differences in fatty acid composition. Finally, ion leakage due to cold-mediated phase separation of the cell membrane could be compared in transformed and non-transformed plants to confirm that expression of the BCOADc gene improves membrane fluidity. Branched chain fatty acids are utilized by many organisms for supporting their growth at low temperatures. This is accomplished by providing the required membrane fluidity at suboptimal temperatures, so that biological functions may take place. Studies over the past three decades showed that it is very difficult to increase the composition of unsaturated fatty acids in membranes of higher plants (Gombos et al. 1994; Moon et al. 1995). Cloning of genes essential for branched chain fatty acid biosynthesis provides a new approach in this research area. In this paper we described the introduction of genes encoding branched chain alpha-oxo acid decarboxylase (BCOADc) into tomato plants. Molecular analysis showed that both the A and B genes were active at transcriptional and translational levels. Comparative analysis of a small number of transgenic tomato plants grown at 4ºC showed that transformed plants had enhanced tolerance to low temperature (4ºC) compared to non-transformed plants. In conclusion, tomato plants transformed with branched chain fatty acid synthesis genes may provide a new model for engineering cold tolerance in plants. In addition to cold tolerance, branched chain fatty acids may contribute to plant development processes such as fruit ripening, senescence, and disease resistance. Delay of fruit ripening in tomato plants has a positive economical impact. Although the introduction of the A and B genes of the BCOADc into tomato plants did not appear to affect the growth morphology, we observed that fruit development in some transformed plants were abnormal. Fruits from plant 1 were smaller and harder. The fruit turned red in similar period of time to the control plants, and the fruit stayed firmer for a much longer period of time. Loss of cell membrane integrity, removal of permeable barriers, and increase in “free space” have been long suggested as major factors in ripening and senescence (Sacher, 1973; Wade and Bishop, 1978; Rouet-Mayer et al. 1995). The role of oxidative breakdown of membrane lipids in plant senescence has attracted new research for many years. It was reported that unsaturated fatty acids are used to synthesize compounds that inhibit the fruit ripening of tomato plants. If the incorporation of branched chain fatty acids can replace unsaturated fatty acids at normal plant growth conditions, the free unsaturated fatty acid pool will be enlarged, resulting in the potential delay of tomato ripening. Acknowledgments We would like to thank Dr. G. Bagi for protein isolation, Mrs. K. Illes for antibody production and Western blot analyses, and Dr. D. Cattani for critical review of the manuscript. Part of the research is the Ph. D. thesis of G. Wang-Pruski completed as University of Alberta, Canada. References
Note: EJB Electronic Journal of Biotechnology is not responsible if on-line references cited on manuscripts are not available any more after the date of publication. Supported by UNESCO / MIRCEN network © 2002 by Universidad Católica de Valparaíso -- Chile The following images related to this document are available:Photo images[ej02025f3.jpg] [ej02025f1.jpg] [ej02025f4.jpg] [ej02025f2.jpg] [ej02025t1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}