 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Electronic Journal of Biotechnology, Vol. 6, No 1., April 15, 2003 REVIEW ARTICLE RNA interference revolution Archana Thakur National Centre for Plant Genome Research, Jawaharlal Nehru University Campus, New Delhi-110067, India Tel: 91 11 6103378 E-mail: a_thakur@hotmail.com Received October 4, 2002 / Accepted March 13, 2003 Code Number: ej03007 RNA interference (RNAi) is a potent method using only a few double stranded RNA (dsRNA) molecule per cell to silence the expression which has made it one of the hottest topics in molecular biology in last two years. Not long ago scientists conducted gene knock out using antisense, dominant negative or knockout techniques which were ineffective, but the discovery of RNAi has enabled to knock out gene in any organism efficiently. RNA silencing was first discovered in transgenic plants, where it was termed co-suppression or post transcriptional gene silencing (PTGS). Five years ago the evidence for RNA silencing emerged from experimental observation on Caenorhabditis elegans, this new approach for achieving efficient, targeted gene silencing is now making its way beyond laboratory notebooks into biotech strategic plans. It’s a phenomenon of gene silencing which offers a quick and easy way to determine the function of a gene. It’s a natural catalytic process and an intrinsic property of every cell of every multicellular organism. Reported to be 1,000-fold more effective than antisense. The new field of RNAi based genomics is increasingly being qualified as a fundamental paradigm shift for biomedical research and development and quite possibly the start of a veritable revolution for the development of modern therapeutics. Keywords: co-suppression, dicer, PTGS, quelling, RISC. DefinitionRNA silencing is a sequence specific RNA degradation process that is triggered by the formation of double stranded RNA that can be introduced by virus or transgenes. Duplexes 21- nucleotide (nt) RNAs with symmetric 2-nt 3’overhangs are introduced into the cell mediating the degradation of mRNA. According to central dogma of molecular biology, proteins are made in two steps. The first step, transcription, copies genes from double stranded deoxyribonucleic acid (ds DNA) molecules to mobile, single- stranded ribonucleic acid (RNA) molecules called mRNA. In the second step, translation, the mRNA is converted to its functional protein form. Since there are two steps to making a protein, there are two ways of preventing one from being made. Scientists have made exciting progress in blocking the protein synthesis through the second step, translation. One way they have accomplished this is by inserting synthetic molecules that triggers a cellular process called RNA interference. DiscoveryMolecular biologists had the dream to knockout gene expression at the mRNA level for the last 15 years. Efforts were made to generate loss of cell functions or organisms various molecules that included from eg antisense sequences, ribozymes and chimeric oligonucleotides. Alternative methods for silencing specific genes have also provided potentially powerful approaches. Antisense methods, using either DNA or RNA, are relatively straightforward techniques for probing gene functions; however, these methodologies have suffered setbacks because of lack of specificity and incomplete efficiency. Moreover, the desired effects were difficult to predict and often only a weak suppression was achieved (Guru, 2000). PTGS in plants involves down-regulation of gene expression at the post transcriptional level, by targeting specific RNAs for degradation. Transgenes are subject to suppression by PTGS, as are other genes that share significant sequence homology with the silenced genes. Plant PTGS is similar to other eukaryotic evidence such as suppression of transgenes and transposons, and cellular responses to double-stranded RNAs. First described in worms in 1998, RNAi operates in plants, fungi, flies and mammals (Fire et al. 1998). Long molecules of double stranded RNA (dsRNA) trigger the process. The dsRNA comes from virus and transposon activity in natural RNAi process, while it can be injected in the cells in experimental processes (Elbashir et al. 2001a). The strand of the dsRNA that is identical in sequence to a region in target mRNA molecule is called the sense strand, and the other strand which is complimentary is termed the antisense strand. An enzyme complex called DICER in D. melanogaster, thought to be similar to RNAase III then recognizes ds RNA, and cuts it into roughly 22- nucleotide long fragments. These fragments termed siRNAs for “small interfering RNAs” which remain in double stranded duplexes with very short 3’ overhangs (Elbashir et al. 2001b) then act as templates for the RNAi inducing silencing complex to destroy the homologous message, thus specifically suppressing its expression. This form of RNAi is termed as PTGS, other forms are also thought to operate at the genomic or transcriptional level in some organisms. Table 1. Examples of RNAi in several species.
The future impact of PTGS is immense that it has a role in viral defence and transposon silencing mechanism. Quelling in fungiIn fungi Neurospora crassa, it was shown that an overexpressed transgene can also induce gene silencing at the post transcriptional level, a phenomenon referred to as “quelling” (Ruvkun, 2001). The first major clue as to the presence of a new mechanism for targeted gene silencing came from an antisense study published in the nematode C. elegans, was reported not only using the expected antisense RNA, but surprisingly, also with the corresponding sense strand. The mystery was solved in 1998, when it was demonstrated that injection of a dsRNA- a common contaminant of many single stranded RNA (ss RNA) preparations yielded a more potent silencing effect than either corresponding ssRNA alone. The process of RNAi has since been recognized as a dsRNA response pathway that has evolved in organisms from plants to human as a natural defense mechanism against molecular parasites such as transposons and RNA viruses. Other manifestations of RNAi had previously been noted as post transcriptional gene silencing in plants and “quelling” in certain fungi. Table 1 elicits the phenomenon of this powerful gene silencing mechanism has been shown to operate in several species among most phylogenetic phyla. Salient features of RNAi
Sequence and strand specific recognitionRNAi is a natural phenomenon believed to occur in the nematode C. elegans, in the fruit fly Drosophila, and in some plant species (Elmayan et al. 1998). It most likely serves to protect organisms from viruses, and suppress the activity of transposons, segments of DNA that can move from one location to another, sometimes causing abnormal gene products. Recent research has shown that an intermediate in the RNAi process, called short-interfering RNAs (siRNA), might be effective in degrading mRNA in mammalian cells. This silencing is gene specific and tissues manifesting degradation have siRNAs complementary to both gene strands. siRNA therefore carries the potential to specifically degrade mRNA that corresponds to mutant genes involved in disease, shutting off the harmful effects of the proteins they encode. RNAi is a potent method, requiring only a few molecules of dsRNA per cell to silence the expression. Not only can silencing spread from the digestive tract of worms to the remainder of the organism, but the effect can also be transmitted through the germ line for several generations.
The natural function of PTGS is providing resistance to virus infection. It has been found tat numerous viruses are potent inducers of PTGS and viruses encode factors that inhibit this response in plants. Systemic nature of PTGS is interesting because it serves an effective way of limiting virus infection to the place of original virus attack (Voinnet and Baulcomb, 1997). PTGS, which was initially considered a bizarre phenomenon limited to Petunias and few other plant species, is now the hottest topics in molecular biology (Cogoni and Macino, 1999). In the last few years it has become evident that PTGS occurs in both plants and animals and has roles in viral defence and transposon silencing mechanisms. Perhaps, the more exciting is the emerging use of PTGS and in particular, RNA interference (RNAi) – PTGS initiated by the introduction of double stranded RNA (ds RNA)- as a tool to knock out expression of specific genes in a variety of organisms (Table 2). These discoveries have intrigued the biological community- an interest that is also fuelled by the increasing power of RNAi as a tool to convert the accumulating hordes of genomic information into a deeper understanding of biological function. Co suppression and PTGS in plants With the development of tools for introduction of transgenes in plants, attempts were made to engineer plants with most desirable characteristics. While trying to increase the purple colour in Petunias, Rich Jorgensen and colleagues (Jorgensen et al. 1994) introduced a pigment producing gene under the control of a powerful promoter. Instead of expected deep purple colour, many of the flowers appeared variegated or even white. This observed phenomenon was named “cosuppression” since the expression of both the introduced gene and homologus endogenous gene was suppressed (Napoli et al. 1990). There are two mechanism through which silencing of both transgenes and endogenous loci could occur. In one mechanism transgene induced silencing was accompanied by heavy methylation of silenced loci, leading to transcriptional gene silencing. In the other mechanism PTGS occurred. This was shown by nuclear run on assays, which indicated transcripts was made but that it failed to accumulate in cytoplasm. Transgene cosuppression is not only limited to plants but has also been shown in fungi, Drosophila, C. elegans and rodent fibroblasts. This phenomenon has been well characterized in Neurospora crassa, where it is known as “quelling” and it occurs at the post-transcriptional level. Although transgene induced silencing in some plants appear to involve gene-specific methylation (transcriptional gene silencing or TGS), in others silencing takes place at the post transcriptional level (post transcriptional gene silencing or PTGS). Mechanism of RNAi – Current working modelsIntensive research efforts to understand this intriguing process elucidates the exact molecular mechanism. In the initiation step the “trigger” ds RNA molecule, usually several hundred base pair long, is cleaved to form 21-23 bp double stranded fragments known as short interfering RNAs (siRNAs) or guide RNAs. siRNAs are produced when the enzyme Dicer a member of the RNAase III family of dsRNS- specific ribonucleases, processively cleaves dsRNA in ATP dependent, processive manner. In the effector step the duplex siRNA are then unwound by a helicase activity associated with a distinct multiprotein complex known as the RNA-induced silencing complex or RISC. An ATP dependent unwinding of siRNA duplex is required for activation of RISC. The siRNA strand that is complementary to the targeted mRNA is then used as primer by an RNA-dependent RNA polymerase (RdRP) to convert the cognate mRNA into dsRNA itself. This dsRNA form of mRNA then becomes a substrate for Dicer cleavage activity, which leads to the destruction of the mRNA and formation of new siRNAs. Effectively, this step amplifies the RNAi response and creates a self- perpetuating cycle of “degradative polymerase chain reaction” that will persist until no target mRNAs remain. This basic ‘core’ pathway defines the RNAi response as one of the most elegant and efficient biochemical mechanisms in nature (Hutvagner and Zamore, 2002). Two C. elegans genes, rdeI and rde4 (rde stands for ‘ RNAi deficient’) are believed to be involved in the initiation step of RNAi. The C. elegans rde1 gene is a member of a large family of genes and is homologous to the Neurospora qde2 (qde stands for “quelling deficient”) and Arabidopsis AGO1 genes (AGO stands for agronaute, AGO1 was previously identified to be involved in Arabidopsis development). Although the functions of these genes is not clear, a mammalian member of RDE1 family has been identified as a translation initiation factor (Sharp, 2001; Silhavy et al. 2002). RNAi effectorsImportant genes for the effector step of PTGS in C. elegans are rde2 and mut7 genes. These genes were initially identified from heterozygous mutant worms that were unable to transmit RNAi to their homozygous offsprings. Worms with mutated rde2 or mut 7 genes show defective RNAi. mut-7 gene encodes a protein with homology to the nuclease domains of RNAase D and a protein implicated in Werner syndrome (a rapid ageing disease in humans) (Grishok et al. 2000). RNA dependent RNA polymeraseNeurospora qde-1, Arabidopsis SDE-1/SGS-2 and C. elegans ego-1, appear to encode RNA dependent RNA polymerase (RdRPs). It might be assumed that this is a proof that an RdRp activity is required for RNAi. Certainly the existence of an RdRp might explain the remarkable efficiency of dsRNA induced silencing if it amplified either the dsRNA prior to cleavage or the siRNAs directly. In C. elegans ego-1 mutants (ego stands for ‘enhancer of glp-1), RNAi functions normally in somatic cells, but is defective in germline cells where ego-1 is primarily expressed. In Arabidopsis SDE-1/SGS-2 mutants (SGS stands for suppressor of gene silencing), siRNA are produced when dsRNA is introduced via an endogenously replicating RNA virus, but not introduced by a transgene. It has been proposed that perhaps the viral RdRP is substituting for the Arabidopsis enzyme in these mutants. Random degradative PCR model suggests that an RdRP uses the guide strand of an siRNA as a primer for the target mRNA, generating a dsRNA substrate for Dicer and thus more siRNAs. Silencing genesSeveral genes controlling RNA silencing in plants have been identified through genetic screens of Arabidopsis mutants impaired in transgene induced RNA silencing (Table 3). They encode a putative RNA-dependent RNA polymerase (SGS2/SDE1), a coiled coil protein (SGS3), a protein containing PAZ and Piwi domains (AGO1) and an RNA helicase (SDE3).The putative SGS2/SDE1 is related to QDE-1 of Neurospora and EGO-1 of C. elegans, and PAZ/Piwi protein AGO is related to QDE-2 of Neurospora, RDE-1of C. elegans, RNA helicase SDE3 is related to SMG-2 of C. elegans and Mut-6 of Chlamydomonas. MUT-7 gene of C. elegans encodes a protein similar to Rnase D, whereas the Drosophila DICER gene encode a protein similar to RNase III. An Arabidopsis ortholog of DICER genehas been identified called CAF, SIN1, SUS1. Developmental role of RNAiAgo1, caf/sin1/sus1 genes in Arabidopsis, ego-1, mut-7 in C. elegans play independent roles in development and silencing. ago1 alleles show normal development but that are deficient in RNA silencing as a null mutant supports the idea that AGO1 participates independently in silencing and development. Plants expressing high levels of viral silencing suppressor HC-Pro or that overexpresses the endogenous suppressor rgs-CAM also exhibit abnormal development. This elicits the possibility that the suppression of RNA silencing mediated via these genes work with interaction of factors that have a dual role in silencing and development. Thus development is interconnected in the pathways of RNAi. Table 3. Genes involved in RNAi.
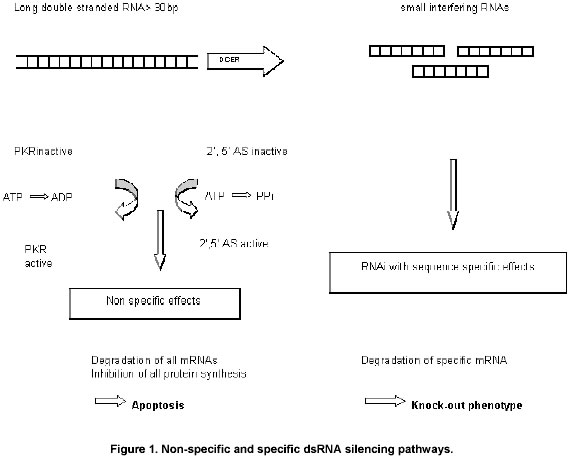
Recent discoveries of small regulatory RNA molecules have identified three types tiny regulatory RNA molecules. Small interfering RNAs (siRNAs) have an integral role in the phenomenon of RNA interference (RNAi). In RNAi, dsRNAs introduced into certain organisms or cells are degraded into ~22nt fragments. These 22nt siRNA molecules then bind to the complementary portion of their target mRNA and tag it for degradation. SiRNAs are believed to have a role in conferring viral resistance and in preventing transposon hopping. The second group of regulatory small RNAs have been referred to as small temporal RNAs. ~22nt lin-4 and let-7 RNAs are example of this group. These RNA molecules have a role in temporal regulation of C. elegans development. These are initially processed from a ~70nt ssRNA transcript folded into a stem loop structure. After processing, these stRNAs are thought to prevent translation of their target mRNAs by binding to the targets complementary 3’ untranslated regions (UTRs). Dicer, RNAase enzyme processes both the types of RNAs (Grishok et al. 2001). Much like lin-4 and let-7, additional small~22nt RNA molecules termed as microRNAs (miRNAs) were discovered in Drosophila, C. elegans and He La cells (Lau et al. 2001; Lee and Ambrose, 2001; Lee et al. 2002; Sharp and Zamore, 2000). These are formed from precursor RNA molecules that fold into a stem-loop secondary structure. MiRNAs play role in regulation of gene expression (Grosshans and Slack, 2002). 100 new mRNAs were identified of which 15% were conserved (with 1-2 mismatches) across worm, fly and mammalian genomes (Sharp and Zamore, 2000). All the identified miRNAs were located at either the 3’ or the 5’ side of a stem loop within a ~70nt RNA precursor. The expression pattern of the miRNAs varied. While some C. elegans and Drosophila miRNAs were expressed in all cells and at all developmental stages, others had a more restricted spatial and temporal expression pattern. This suggested that these miRNAs might be involved in post transcriptional regulation of developmental genes. Systemic silencing in plantsThe mechanism of PTGS is initiated from dsRNAs that result from replicative intermediates of viral RNAs or aberrant transgene coded RNAs. PTGS that starts locally in plants by a transgene or virus can spread systematically to the rest of the plant. dsRNAs or siRNAs when introduced locally in the plants can trigger systemic silencing. siRNAs act as “mobile trigger elements” for systemic silencing. A remarkable feature of RNA silencing is its ability to act beyond the cells in which it is initiated. In plants carrying an expressed GFP transgene, RNA silencing can be initiated by localized introduction of an additional ectopic GFP transgene. The RNA silencing is initially manifested in the tissues containing the ectopic DNA but eventually becomes systemic showing that a silencing signal moves between cells and in the vascular system of the plants. Animals also have a system for amplification and spread of silencing. This is quite evident in C. elegans. If these animals are injected with dsRNA or if they are allowed to feed on bacteria that produce dsRNA there is systemic silencing of a corresponding endogenous RNA. It is presumed that the signal molecule is produced in the C. elegans cells that receive dsRNA. This signal then moves and causes RNA silencing in other cells that become sources of secondary signal. PKR is responsive protein kinase and the cascade of activation and inactivation induces a global non-specific suppression of translation which in turn triggers apoptosis (Williams, 1997). Application of RNAi in mammals: Non-specific and specific dsRNA silencing pathways (Figure 1) The presence of extremely low levels of viral dsRNA triggers an interferon response (called acute-phase response) and the activation of a dsRNA responsive protein kinase (PKR). PKR phosphorylates and inactivates translation factor EIF-2a leading to activation of the 2’,5’ oligo adenylate synthetase, finally resulting in RNAase L activation. This cascade induces a global non-specific suppression of translation, which in turn triggers apoptosis (Xu et al. 2001; Gill and Esteban, 2000). Small dsRNA called siRNA specifically switched off genes in human cells without initiating the acute phase response. Thus these siRNAs are suitable for gene target validation and therapeutic applications in many species, including humans. The recent success in triggering the RNAi pathway in vertebrate systems now opens the door to direct use of dsRNA molecules as therapeutic agents with exquisitely controllable specificity to alleviate human disease. In addition to the promise of finally achieving truly personalized machines, this approach holds the potential for greatly accelerated and more cost-effective preclinical development while bypassing some of the key obstacles met by antisense therapeutics, for example, the instability of ssRNA molecule. RNAi and antisense inhibitionAntisense methods, using either DNA or RNA, are straight forward techniques for probing gene functions, however the discrepancy lies in the fact that this process suffers from specificity and incomplete efficacy. RNA silencing is induced in plants at varying efficacies by transgenes designed to produce either sense or antisense transcripts. Furthermore, transgenes engineered to roduce self complementary transcripts (dsRNA) are potent and consistent inducers of RNA silencing. Short- interfering RNA could provide medical researchers new hope in using gene silencing for therapeutic purposes. Until now, another gene silencing technique, using antisense oligonucleotides, had been the main hope for clinical application. Antisense oligonucleotides are short pieces of DNA or RNA complementary to sequences on RNA. They are believed to work by hybridizing to the mRNA, creating a double stranded stretch, which slows down ribosome transcription. Antisense DNA creates RNA-DNA duplexes that are most likely recognized by RNAase H, an enzyme that cuts double stranded molecules containing one DNA and one RNA strand, which cleaves the mRNA (Crooke, 1999). Since the mRNA is cut, it cannot be translated into a functional protein product. While this process can be triggered somewhat effectively in vitro, antisense technology has not completely lived up to expectations due to difficulties in delivering oligonucleotides to cells in vivo and problems with the accessibility of specific sites on mRNA (Caplen et al. 2001). The RNAi distinguishes itself markedly from the much less potent process of antisense inhibition both by its enzymatic nature and by its inherent irreversibility of its mode of action, i.e. destruction of the targeted mRNA. Also, by relying on the use of dsRNA molecules it largely avoids the major problem of ssRNA instability that has long plagued the antisense field. RNAi has also proved far superior to ribozyme based paradigms by showing equal potency in vivo as it does in vitro. Finally the RNAi response also exhibits extremely stringent sequence specificity- a single base pair mismatch over the length of an siRNA is enough to block the response. Already from these basic facts, RNAi clearly emerges as the ideal method for achieving targeted gene silencing in vivo. It has more potential for success since it seems to be more stable than single stranded antisense molecules, making cellular delivery easier. So far all the siRNAs tested in mammalian cells have inhibited expression of the target genes, which is an encouraging sign (Caplen et al. 2001). If siRNA can indeed be an effective silencer, it could serve many important uses in functional genomics and to silence oncogenes. Antisense molecules have already been used in cancer therapies (Yu et al. 2002) but it is hard to get them into cells and to cleave their target efficiently. Perhaps the siRNA stability will prove to make it more efficient at getting to and eliminating its target than the antisense oligonucleotides tried so far. Functional genomics and RNAiRNA silencing is well suited to the systematic analysis of gene function. This principle is simple: a gene fragment is introduced into a cell as dsRNA or as DNA that will give rise to dsRNA. The dsRNA activates the DICER/RISC process so that the properties of the affected cell reflect a loss of function in the corresponding gene. In order to target the genome of C. elegans a collection of E. coli strains have been generated to produce dsRNA corresponding to each gene. The function of each gene is inferred from the behavior or properties of the C. elegans after feeding on the E. coli. A parallel programme is been carried out in plants infected with viruses containing inserts corresponding to each of the genes in the plant genome. The function of the gene can be inferred from the symptoms that develop in the infected plant. Recently a number of groups have developed expression vectors to continually express siRNAs in transiently and stably transfected mammalian cells (Lee et al. 2002; Sui et al. 2002). Some of these vectors have been engineered to express small hairpin RNAs (shRNAs), which get processed in vivo into siRNAs like molecules capable of carrying out gene specific silencing (Ruiz et al. 1998; Lohmann et al. 1999; Brummelkamp et al. 2002; Yu et al. 2002). Silencing genes in HIVThe strategy used was to silence the main structural protein in the virus, p24, and the human protein CD4, which the virus needs to enter the cells. This impairs the virus in infected cells and limits its spread into healthy cells (Paddison et al. 2002). Hence the production of virus is inhibited either by blocking new infections or blocking the production of new viral particles in infected cells (Lohmann et al. 1999). The concept of silencing genes in HIV is straightforward: Hit the virus where it counts by eliminating a protein it needs to reproduce or cause infection. siRNA molecules (shorter than 30 base pairs) are added to the cells, where the cell recognizes and degrades mRNA corresponding to the target sequence. As a result little or no protein is produced. What makes RNAi so exciting to the researchers is its potential for knocking out a protein without harming a cell. By comparison chemotherapy kills tumors by destroying cancerous as well as healthy cells. RNAi strategy includes multiple targets to kill HIV. These could be the targets that block entry into the cells and disrupts the virus life cycle inside the cells. This technology will help researchers dissect the biology of HIV infection and design drugs based on the information. Researchers at City of Hope Cancer Centre in Duarte have developed a DNA-based delivery system in which human cells are generated that produce siRNA against REV protein, which is important in causing human disease (Yu et al. 2002). Transgenic tomatoes shut the disease invading geneTransgenic tomatoes have been generated which are resistant to crown gall disease. The plants were engineered to contain versions of two bacterial genes required to cause the disease. This is the first report of control of a major bacterial disease through gene silencing. The extra genes recognize and effectively shut down the expression of the corresponding bacterial genes during infection, thus preventing the spread of infection. The incoming bacteria cannot make the hormones they need to make the tumors. Crown gall disease is a perennial problem for growers of ornamental plants, such as roses, fruits and nuts. Virus induced gene silencing (VIGS)Replication of plant viruses, many of which produce dsRNA replication intermediates, very effectively cause a type of RNA silencing called VIGS (Virus induced gene silencing).When viruses or transgenes are introduced into plants, they trigger a post transcriptional gene silencing response in which double stranded RNA molecules, which may be generated by replicative intermediates of viral RNAs or by aberrant transgene coded RNAs. The dsRNAs are then digested into 21-25 nt small interfering RNAs or siRNAs. The siRNAs subsequently assemble into a nuclease complex called RISC, guiding the complex to bind and destroy homologous transcripts. PTGS is believed to be an anti-viral response. Viral RNAs not only trigger PTGS, but they also serve as targets. Cleavage of viral RNA results in reduce virus titers in local and distant leaves and a plant recovery phenotype.In response numerous plant viruses have evolved proteins to suppress PTGS (Elbashir et al. 2001a). One of these suppressor proteins, the tombavirus p19, abolishes gene silencing by binding to 21-25nt siRNAs (Wang et al. 2000) could suppress PTGS in Nicotiana benthamiana plants. Coinfilteration with green fluorescent proteins and p19 results in retention of the green fluorescence in plant leaves. GFP/p19 coinfilteration (i.e. suppressed PTGS) resulted in increase GFP mRNA levels and decreased GFP siRNA levels in plant leaves compared to GFP infiltration alone. The results by Silhavy and his colleagues (Sui et al. 2002) suggests that tombavirus p19 protein suppresses local PTGS by binding to 21-25nt siRNAs. Inhibition of local silencing may be achieved by preventing siRNAs from interacting with either RdRp enzyme or the RISC complex. In addition to local silencing p19 could prevent systemic silencing. In systemic silencing, infilteration of plants with GFP allows PTGS to spread to distance leaves. It has been determined that the spread of PTGS does not involve movement of the Agrobacterium tumefaciens bacteria or the Ti plasmid. Rather it has been proposed that this ‘mobile silencing element’ may either be long dsRNAs or siRNAs (Voinnet et al. 2000; Wang et al. 2000) Therefore siRNAs and not the longer dsRNAs, act as mobile silencing element since p19 can inhibit systemic silencing and p19 can only bind to siRNAs. In addition to suppressing PTGS in plants, p19 may also be used as a tool to inhibit RNAi in other systems. This is suggested by the finding that p19 can bind to siRNAs in vitro without requiring host proteins or other factors. Epigenetic control – RNA based mechanism Epigenetic control of gene expression can be considered from the stand point of normal development which is accomplished by the stable and visible repression of specific genes. Sequence specific RNA degradation: A homology based PTGS process that targets specific RNAs for degradation has been discovered in diverse organisms. Each involves the induction of sequence –specific RNA turnover by the introduction of several hundred bases of RNA sequence present in the mature target mRNA. Two remarkable features of PTGS are amplification and cell to cell movement of sequence specific signal. Amplification is more apparent in C. elegans, where the concentration of dsRNA required to initiate turnover is much lower than the concentration of target RNA. Amplification or some type of relay system also operates in plants. Cell to cell movement of the silencing agent has been observed in plants and C. elegans. In plants, long distance transport occurs through the vascular system and is thought to involve a mobile silencing signal containing dsRNA. RNA directed DNA methylation: Nuclear and cytoplasmic processes can be connected through dsRNA molecules. A second thought is possibly the transcription of aberrant RNAs from methylated DNA templates. These aberrant RNAs have been postulated to trigger RNA turnover in the cytoplasm and methylation of unlinked homologous DNA copies. The ability of RNAs produced in the cytoplasm to feedback and induce epigenetic changes on DNA has been most clearly shown in plants, where nuclear transgenes became methylated only subsequent to the initiation of transgene PTGS in the cytoplasm by an RNA virus engineered with transgene sequences. dsRNAs are actual inducer of DNA methylation has been suggested by the ability of viroids, a plant pathogen consisting solely of a non coding RNA duplex to trigger methylation of homologous nuclear DNA. Applications and future of RNAi technologyIn context with the current status of knowledge about RNAi, it is a revolution in the field of plant molecular genetics that it has enormous potential for engineering control of gene expression, as well as for the use of a tool in functional genomics. The ability to manipulate RNA silencing has a wide variety of practical applications of biotechnology ranging from molecular biology to gene therapy in animals. This process can be induced experimentally with high efficiency and targeted to a single specific gene or a multigene family. The use of RNAi as a method to alter gene expression has been attempted in a diverse group of organisms, employing different methods, with different rates of success. In C. elegans, Drosophila and plants, RNAi seems to be an effective, specific and valuable tool for reverse genetics. A second group including zebrafish, Xenopus and mouse show RNAi with some limitations. RNA interference employing short dsRNA oligonucleotides will permit to decipher the functions of genes being only partially sequenced. One of the first commercial products of RNA silencing was tomato in which the target was to reduce the expression of these genes in the silenced plants meant that the tomatoes were firm after ripening and were not damaged by handling. Virus induced gene silencing is potentially a powerful tool to silence the endogenous genes that are homologus to any sequences carried within the virus. This technology will enable to use plant virus induced gene silencing approach for plant genetic studies. RNAi is important for inhibition of gene expression at the post transcriptional level in eukaryotic cells. Worms can be engineered for the generation of stable phenotypic null mutants. In this context RNAi is a straight forward tool to rapidly assess gene function and reveal null phenotypes. Development of RNAi technology for the use in post implantation embryos (Brown et al. 2002). Drosophila embryology studies reveal to the production of null phenotypes by injecting early stage embryos. RNAi technology can be applied as genetic tools in vertebrates to induce sequence specific silencing in early mouse embryos. The predominant economic significance of RNA interference is established by its application as a therapeutic principle. As so, RNAi may yield RNA based drugs to treat human diseases (Jarvis and Ford, 2001). siRNA is effective against parasites, so perhaps it can be used to silence parasitic genes (Fire, 1999) or used against other pathogens to benefit host organisms like humans (Ruiz et al. 1998). In most mammalian cells, 400-500 base pair long dsRNA invokes a more general translational suppression through a pathway involving interferon, ultimately leading to cell death by apoptosis. (Romano and Macino, 1992). dsRNA can silence the expression of exogenous genes in Chinese hamster ovary cells. Hence, siRNA brings the possibility of specific gene silencing through mRNA degradation, something its precursor, dsRNA, cannot do, while possibly being more versatile than less stable single stranded antisense oligonucleotides.
Note: Electronic Journal of Biotechnology is not responsible if on-line references cited on manuscripts are not available any more after the date of publication. Supported by UNESCO / MIRCEN network © 2003 by Universidad Católica de Valparaíso -- Chile The following images related to this document are available:Photo images[ej03007f1.jpg] |
| |||||||||

{kind=link}