 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Electronic Journal of Biotechnology, Vol. 6 No. 2, August 15, 2003 SHORT COMMUNICATION Infection of Caenorhabditis elegans by Salmonella typhi Ty2 Javier Santander1, Juan Carlos Espinoza2, María Soledad Campano3, James Robeson*4 1Laboratorio de Bacteriología,
Instituto de Biología,
Pontificia Universidad Católica de Valparaíso,
Av. Brasil 2950, Valparaíso, Chile
Tel: 56 32 273118
Fax 56 32 273109
E-mail: jrobeson@ucv.cl Financial support: Vice-Rectoria de Investigación y Estudios Avanzados, Universidad Católica de Valparaíso. Proyecto D.I.UCV 122.770.2002. Received January 23, 2003 / Accepted July 15, 2003 Code Number: ej03018 Abstract Several serovars of Salmonella infect and kill the nematode C. elegans. However, here we report that Salmonella typhi Ty2, a representative strain of this human pathogen, readily infects the intestinal lining of C. elegans without significantly affecting its viability. Our observation suggests extending the use of the C. elegans model system for the study of host parasite relationships, to address problems concerning the biology of S. typhi. Keywords: Caenorhabditis elegans, invasiveness, Salmonella typhi Ty2. Article The nematode Caenorhabditis elegans has been used as a model system to study bacterial pathogenesis due to ease of manipulation and a detailed knowledge of its biology. Several bacterial pathogens, both Gram positive and Gram negative, have been reported to infect and kill C. elegans (Couillault and Ewbank, 2002). Recently, C. elegans has been used to elucidate molecular mechanisms of virulence in Pseudomonas aeruginosa (Gallagher and Manoil, 2001) infection by Burkholderia paseudomallei (O’Quinn et al. 2001) and S. typhimurium, a bacterium that persistently infects the C. elegans intestine and finally kills the nematode (Aballay et al. 2000; Aballay and Ausubel, 2001). Furthermore S. enteritidis and S. dublin have also been shown to kill C. elegans (Aballay et al. 2000). On the other hand, S. typhi is considered to be a pathogen restricted to human hosts (Pascopella, et al. 1995) and therefore not many cell or animal systems are available to study S. typhi pathogenesis. Here, we report that the S. typhi Ty2 WHO reference strain does not kill C. elegans but can infect the nematode’s intestinal lining. Consequently, C. elegans is suitable for exploring cell invasion by S. typhi and possibly its persistence in this host. Growth of bacteria and C. elegans Both Wild Type (WT) and Green Fluorescent Protein (GFP) tagged bacteria were used. The latter contained the plasmid pSU2007 that codes for GFP and Kanamycin resistance (Kmr). S. typhi Ty2 WT, S. typhi Ty2 pSU2007, S. typhimurium SL1344, S. typhimurium SL1344 pSU2007, Escherichia coli MT102 pSU2007 and E. coli OP50 were grown in Luria-Bertani medium (Miller, 1972) at 37ºC. The nematode C. elegans WT N2 Bristol was propagated on NG agar, fed with E. coli OP50 (Brenner, 1974). Assays were performed according to Aballay
et al. 2000. Dead nematodes were counted every 24 hrs. and removed from
the assay plates. Thus, we determined the time it takes for 50% of the nematodes
to die (TD50).
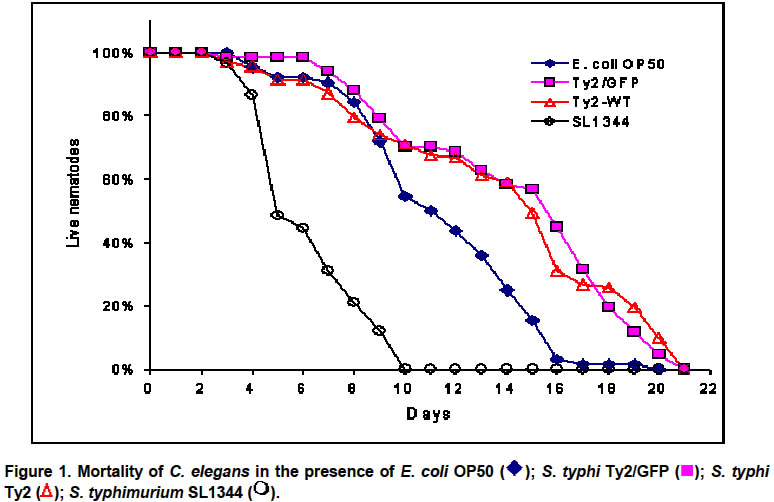
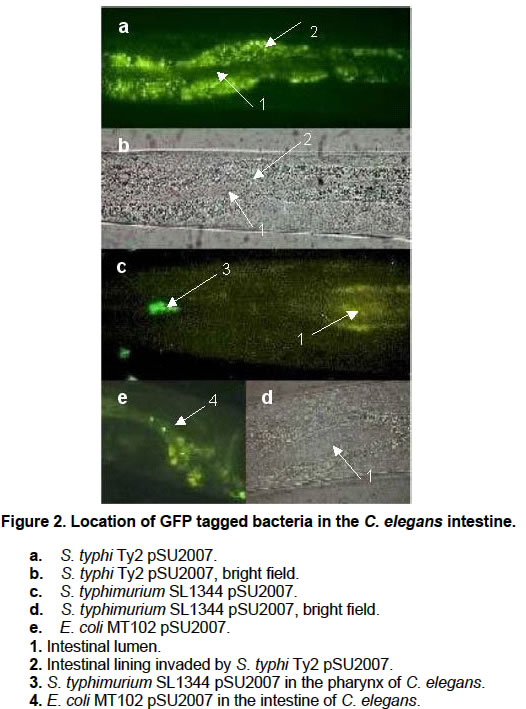
Nematodes infected with different GFP-tagged bacteria, were suspended in M9 salts solution (Miller, 1972) for 10 min., centrifuged and finally suspended in M9 with 30 mM sodium azide, used as anesthesic (Aballay et al. 2000). After the worms ceased to move they were observed by epifluoresence microscopy at 460-490 nm using a Olympus BX 60 microscope. Images were obtained using an Olympus C3030-Zoom digital camera. A total of 50 specimens were examined, coming from four independent C. elegans – S. typhi Ty2 plates. Results and Discussion Recently, Aballay et al. 2000 have reported a TD50 of 7,6 +/- 0,7 days for a nosocomial isolate of S. typhi (strain 469) in a 10 day experiment designed to assay killing by S. typhimurium SL1344. However, when assaying the WHO reference strain S. typhi Ty2 we found that it does not kill C. elegans in a 22 day assay (Figure 1). We found TD50’s of 14,94 days for S. typhi Ty2 WT, 15,56 days for S. typhi Ty2 pSU2007, 11 days for E. coli OP50 and 4,97 days for S. typhimurium SL1344. No swelling of the intestine that was observed in S. typhi Ty2 infected C. elegans (Figure 2a and Figure 2b) in contrast with S. typhimurium SL1344 infected nematodes (Figure 2c and Figure 2d). In addition, we saw that S. typhi Ty2 invades the worm’s intestinal lining (Figure 2a). This is consistent with a reduced reproductive rate we observed for C. elegans grown in S. typhi Ty2 (48,2 worms/ml/day) when compared to the reproductive rate of E. coli OP50 grown nematodes (96,2 worms/ml/day). These results suggest that nematodes, such as C. elegans, might act as temporal reservoirs for this bacterium. In this respect, Tesser et al. 2001 have reported carriage of S. typhi inside environmental protozoa, which act as potential reservoirs. The fact that C. elegans infected with S. typhi remains viable and active suggests that bacterivorous nematodes might play a role in the dispersal of S. typhi. We are currently testing this possibility in view of recent evidence (Chadfield et al. 2001) indicating that the poultry parasitic nematode Ascaridia galli is involved in the dispersal of S. typhimurium. In this case, the bacterium infects A. galli but does not kill it, thus promoting its own dissemination. Finally, the C. elegans - S. typhi Ty2 association allows to address questions about invasiveness of S. typhi in a whole organism system, with the added advantage of the detailed knowledge pertaining the genetics and molecular biology of C. elegans. This is a complementary approach to a simpler cultured cell system expressing a surface receptor for S. typhi that has been described earlier (Pier et al. 1998). Furthermore, the C. elegans system could be useful in elucidating differences in host specific adaptations between S. typhi and S. typhimurium, considering that the latter remains in the intestinal tract during the lethal infection of C. elegans (Aballay et al. 2000). Acknowledgments We thank Dr. Guido Mora (Pontificia Universidad Católica de Chile) who provided S. typhi Ty2, Dr. Alejandro Aballay (Harvard Medical School) for C. elegans, S. typhimurium SL1344 and E. coli OP50, Dr. Soren Mölin (Molecular Ecology Group, BioCentrum-DTU, Denmark) for E. coli MT102 pSU2007 and Dr. Juan Kuznar (Universidad de Valparaíso) for use of epifluorescence microscopy facilities. References
Note: Electronic Journal of Biotechnology is not responsible if on-line references cited on manuscripts are not available any more after the date of publication. Supported by UNESCO / MIRCEN network © 2003 by Pontificia Universidad Católica de Valparaíso -- Chile The following images related to this document are available:Photo images[ej03018f1.jpg] [ej03018f2.jpg] |
| |||||||||

{kind=link}
{kind=link}