 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Electronic Journal of Biotechnology, Vol. 6 No. 3, December 15, 2003 Therapeutic angiogenesis following intramuscular gene transfer of vascular endothelial growth factor 121 in a dog model of hindlimb ischemia Ariana G. Ojalvo*1, Alina Seralena2, Raysa Vázquez3, José F. Montequín4, Nelson S. Vispo5, Ricardo Silva6, Alfredo Aldama7, Yaquelin Puchades8, Luis T. Sorell9, Pedro Lopez-Saura10, María A. Alfonso11, Rafael Simón (†)12, Alfonso Alí13, Armando Seuc141, Luis Herrera15 1Centro de Ingeniería
Genética y Biotecnología,
Ave. 31 e/ 158 y 190, Cubanacán,
P.O. Box 6162, La Habana 10600, Cuba
Tel: 53 7 271 6022
Fax: 53 7 2714764
E-mail: ariana.garcia@cigb.edu.cu Financial support: This work was supported by the Center for Genetic Engineering and Biotechnology, Havana, Cuba. Received January 31, 2003 / Code Number: ej03025 Abstract Vascular endothelial growth factor (VEGF), an endothelial cell-specific mitogen, has been shown to promote therapeutic angiogenesis in animal models of critical limb ischemia. Ischemic skeletal muscle is advantageous for taking up and expressing foreign genes transferred as naked plasmid DNA. Accordingly, we investigated the hypothesis that intramuscular administration of naked plasmid DNA encoding the 121-amino acid isoform of VEGF could augment collateral development and tissue perfusion in a dog hindlimb ischemia model. Unilateral hindlimb ischemia was surgically induced in Beagle dogs. Ten days later, animals received intramuscular injections of pVEGF121 plasmid directly in the ischemic muscles. Angiogenic effects were evaluated by angiography, calf blood pressure ratio and vasomotor reserve analyses. Thirty days after gene transfer, angiographically recognizable collateral vessels were increased in pVEGF121-treated animals compared with controls. Improvement in perfusion to the ischemic limb was documented by a significantly higher calf blood pressure ratio for pVEGF121 (0.79 ± 0.05) versus controls (0.56 ± 0.14, P<0.01). Vasomotor reserve assay suggested amelioration in blood availability at the microcirculation level in pVEGF121-treated animals. Hematological variables showed no significant modification due to the treatment. Our results suggest that intramuscular gene transfer of VEGF121 may promote therapeutic angiogenesis in critical limb vascular insufficiency. Keywords: collateral development, gene therapy, naked plasmid DNA, peripheral vascular disease, revascularization.
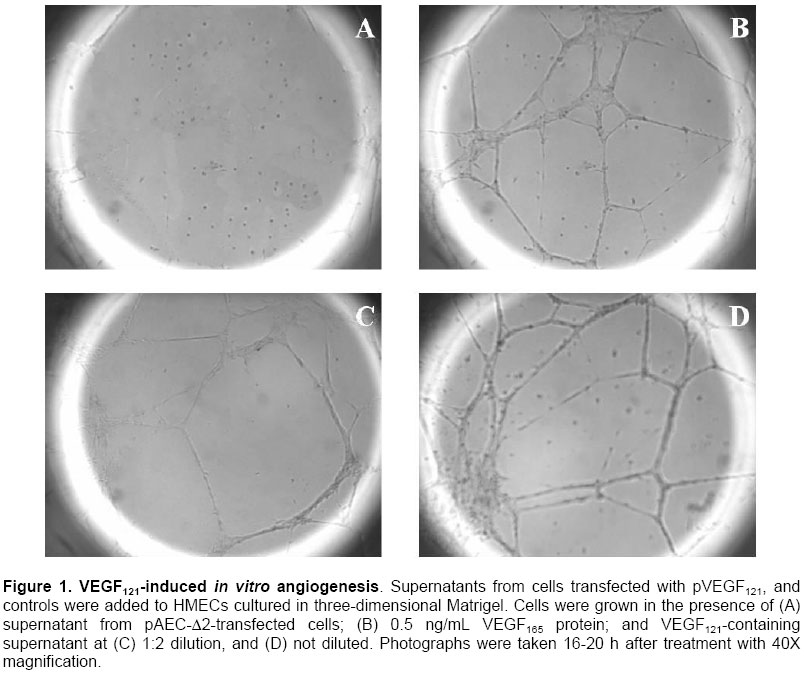
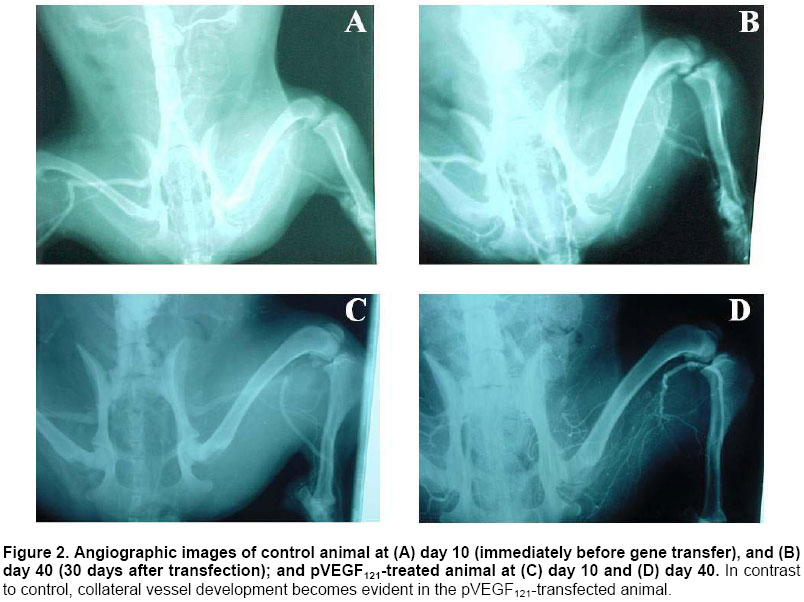
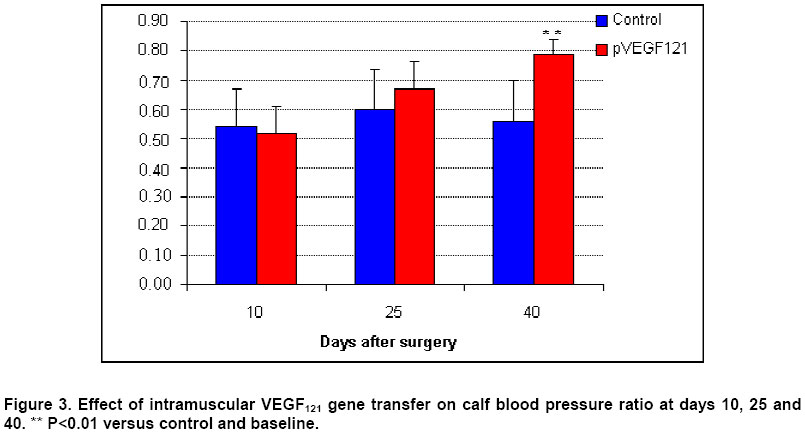
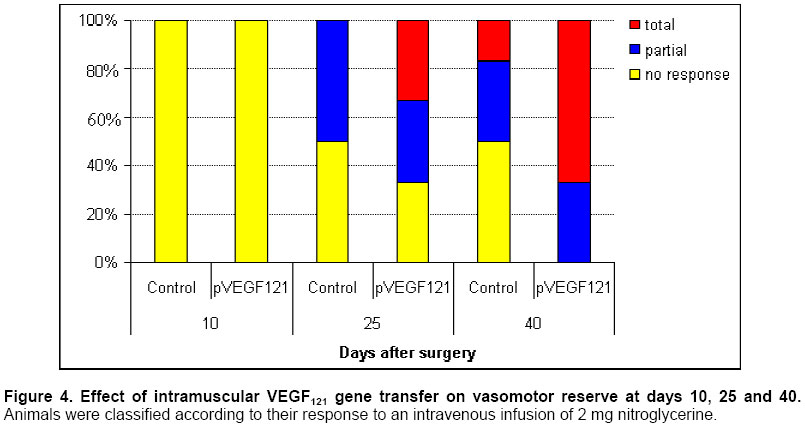
Critical limb ischemia is estimated to develop in ~500 to 1000 individuals per million per year (Creager, 2001). Conventional drug therapy is of no proven benefit for these patients (Nikol and Huehns, 2001). Despite major advances in both percutaneous and surgical techniques, the disease frequently follows an inexorable down-hill course (Gardner and Killewich, 2001). Indeed, many of these patients face amputation (Dormandy et al. 1999), which is associated to high morbidity and mortality, and patient’s life quality becomes remarkably affected. Consequently, the need for alternative strategies for the treatment of patients with critical limb ischemia is compelling. Therapeutic angiogenesis is a novel concept consisting in the use of angiogenic growth factors to expedite and/or augment collateral artery development in ischemic tissues. It constitutes a potential alternative approach for treating vascular insufficiencies. Vascular endothelial growth factor (VEGF) (Leung et al. 1989), a heparin-binding dimeric glycoprotein, is among the various growth factors that have been shown to promote angiogenesis. VEGF is an endothelial cell specific mitogen (Neufeld et al. 1999) that is upregulated under hypoxia (Semenza, 2001). Preclinical studies using recombinant VEGF have proven its efficacy in animal models of lower limb (Bauters et al. 1995; Walder et al. 1996) and myocardial ischemia (Hariawala et al. 1996; Lopez et al. 1998). However, recombinant protein therapy has several limitations for therapeutic angiogenesis. First, a single or multiple bolus doses of recombinant protein are not effective to maintain an optimally high and local concentration over time. Second, protein therapy requires expensive manufacturing faculties and years of scale-up effort. Third, rhVEGF has been shown to produce systemic hypotension (Hariawala et al. 1996; Horowitz et al. 1997). Gene therapy presents a solution to the dosing dilemma, since it can be considered a sophisticated form of a sustained delivery system. Moreover, gene therapy is cheaper to develop, implement, and reimburse. Striking advantages of VEGF gene transfer in particular are its specificity for the target cells, with minimal transgene expression in other tissues; and its low systemic effect due to a short half life (29-77 min) in the circulation (Eppler et al. 2000). Several gene transfer studies have demonstrated that VEGF expression can stimulate the development of collateral vessels in a rabbit hindlimb ischemia model, using arterial (Takeshita et al. 1996b; Takeshita et al. 1996c) or intramuscular (Tsurumi et al. 1996; Tsurumi et al. 1997) administration. The results of clinical trials (Isner et al. 1996; Baumgartner et al. 1998; Isner et al. 1998) document that gene transfer of naked plasmid DNA encoding VEGF promotes collateral vessel development in patients with critical limb ischemia. Adenovirus-mediated gene transfer of VEGF121 has been shown to improve lower limb flow reserve and endothelial function in patients with peripheral arterial disease (Rajagopalan et al. 2001). Several studies with VEGF121 have used the same approach to promote angiogenesis in the ischemic myocardium of animals (Patel et al. 1999; Lee et al. 2000), and patients (Rosengart et al. 1999a; Rosengart et al. 1999b). However, virus administration can lead to immune responses that could cause serious immunopathology. The use of naked plasmid DNA obviates these immunological concerns and clearly simplifies the transfection protocol. The present study was designed to test the hypothesis that intramuscular administration of naked plasmid DNA encoding the 121-amino acid isoform of VEGF could augment collateral development and tissue perfusion in a dog hindlimb ischemia model. Materials and Methods Cloning VEGF121 gene The DNA sequence encoding human VEGF121 was amplified from the plasmid hVEGF121 (Plate et al. 1992) by polymerase chain reaction (PCR) and ligated into the expression vector pAEC-D2 (Herrera et al. 2000) digested with PvuII and BamHI. This vector contains the immediate-early promoter/enhancer from the human cytomegalovirus (CMV) to drive VEGF121 expression. Downstream from the VEGF121 gene are the SV40t splicing/polyadenylation signals (SV40t SS/pA). The kanamycin resistance marker (transposon 903) is also included in this plasmid. These fragments occur in the pUC19 vector, which includes the Escherichia coli ColE1 origin of replication. The recombinant plasmid was named pVEGF121. Bacterial culture and purification of pVEGF121 E. coli DH10B cells [mcr A, mcr B (a), mrr (b), hsd R17(c), deoR (rec A1) end A1, lac ZD M15 (a) deletion of mcrBC (b) deletion of hsd R and hsd M] transformed with pVEGF121 plasmid were grown in a fermentor containing 5 L of Terrific Broth medium with 50 mg/mL kanamycin at 37ºC, 500 rpm. agitation, and an air flow rate of 5 L/h. For increased copy number, the temperature was raised to 42ºC at mid-log phase (Lahijani et al. 1996). After bacterial alkaline lysis, the mixture was neutralized and clarified by tangential flow filtration (TFF) using a nominal molecular weight cutoff TFF polyethersulfone membrane (1,000,000 Da). Subsequent chromatography purification steps were performed as follows: Q-sepharose (Amersham Pharmacia, UK) fast flow anion exchange chromatography (AEC) and Sephacryl S-1000 (Amersham Pharmacia, UK) gel filtration chromatography. In vitro transfection and expression of VEGF121 CHO-K1 cells (ATCC CCL-61) were transfected using 1 mg pVEGF121 plasmid by the polyethylenimine (PEI) (Aldrich, USA) method. In order to control transfection, cells were co-transfected with 5 mg pAGFP plasmid, carrying the green fluorescent protein (GFP) gene under the control of the CMV promoter. Transient co-expression of VEGF121 and GFP was achieved in D-MEM (Sigma, USA) containing 10% FCS and 5 mg/mL gentamycin. To confirm VEGF121 expression, 48-h culture supernatants from transfected CHO-K1 cells were assayed by ELISA (Quantikine, R&D Systems, USA). Matrigel angiogenic activity assay Terasaki plates (Nunc, USA) were coated with Matrigel basement membrane matrix (Becton Dickinson, USA) (Benelli and Albini, 1999). Approximately 103 human microvascular endothelial cells (HMEC), from E Ades (CDC, USA), were added per well. HMECs were grown for 16-20 h in the presence of 48-h culture supernatants from CHO-K1 cells transfected with pVEGF121. A supernatant from cells transfected with pAEC-D2 vector was used as negative control, and VEGF165 protein (0.5 ng/mL) was used as positive control. Finally, the wells were photographed under the inverted microscope. Beagle dogs with surgically induced unilateral hindlimb ischemia were used for the experiments. All protocols were approved by the Institutional Animal Care and Use Committee and were conducted in accordance with the Health Guide for the Care and Use of Laboratory Animals. Twelve Beagle dogs, weighing 11-13 kg, were anesthetized with thiopental (20 mg/kg). A longitudinal incision was then performed, extending from 3 cm over the crural region to the pre-patellar region of one limb. Through this incision, the femoral artery was dissected free, along its entire length; all branches of the femoral artery, including the inferior epigastric, deep femoral, lateral circumflex, and superficial epigastric arteries, were also dissected free. After further dissecting the popliteal and saphenous arteries distally, the external iliac artery and all of the above arteries were ligated (3.0 silk; Ethicon, Sommerville, NJ). Finally, the femoral artery was completely excised from its proximal origin as a branch of the external iliac artery, to the point distally where it bifurcates to form the saphenous and popliteal arteries. Postoperation, all animals were closely monitored. Subcutaneous penicillin (106 units daily, during 7 days) was administered for prophylaxis. Ten days after surgical induction of ischemia, animals received intramuscular injections with pVEGF121 plasmid (n=6) or placebo (n=6), using 1-mL syringes and 26 G needles. Injections were performed slowly to prevent fluid loss from the epimysium. Each dog was injected at four different sites in the ischemic muscles. One pVEGF121 dose contained 2 mg / 2 mL; each dose was divided in four 0.5 mg / 0.5 mL injections. Placebo consisted of sterile saline (0.9% NaCl), and it was administered in the same way as pVEGF121 (four 0.5 mL injections). After palpation of the aorta beat, a long 18 G needle was introduced into the aortic artery and 20 mL of contrast media (sodium diatrizoate; Chinoin, China) were injected in a single puncture. Images of the ischemic hindlimb were recorded on 11x14 inches films. This procedure was carried out using the aforementioned anesthetic. Angiography was performed on day 10 (baseline), and again on day 40. Ischemic/normal limb blood pressure ratio was measured by infrared photopletismography. Pressures were obtained by placing an appropriate pneumatic cull above the knee-joint of the rear legs. The infrared photosensor was placed on the rear foot. The systolic blood pressure was determined by slowly deflating the cuff pressure from the supra-systolic values and recording the first inflow with the photocell used as a pulse sensor. Vasodilator response was assessed by intravenous infusion of 2 mg nitroglycerine. Systolic amplitude of the peripheral pulse wave was measured before and every five seconds after the nitroglycerine infusion during 10 min. Vasodilator responses were classified as no response, partial (for less than 10 min.) or total (for 10 min. or more). Animals were evaluated according to a set of hematological variables including hemoglobin, hematocrit, glycemia, creatinin, total proteins, albumin, alkaline phosphatase (AP), glutamic-pyruvic transaminase (GPT), glutamic-oxaloacetic transaminase (GOT), coagulation time, bleeding time, and platelet counting. Measurements were performed at days 10 (baseline), 20 and 40. Calf blood pressure ratio results were expressed as mean ± SD. The Shapiro-Wilk test was used to analyze normal distribution. Statistical significance was evaluated using a Student’s t test for equal variances, two-tailed distribution. To analyze vasomotor reserve experiment, a Fisher’s exact test was used. Data from hematological variables were expressed as mean ± SD, and evaluated by the Mann Whitney’s U test. Values of P<0.05 and P<0.01 were interpreted to denote statistically significant (*) and very significant (**) differences, respectively. Cloning VEGF121 gene The human VEGF121 gene was inserted into the expression vector pAEC-D2 (Herrera et al. 2000). The resultant plasmid, named pVEGF121, with a size of 4039 bp, contains the VEGF121 gene under control of the immediate-early promoter/enhancer from the human CMV. The kanamycin resistance gene was used for appropriate selection in bacterial systems without undesirable side effects in gene therapy. The insert was confirmed by restriction analysis and DNA sequencing (data not shown). A process combining alkaline lysis, TFF, anion exchange and size exclusion chromatography, yielded a 99% pure, >95% supercoiled plasmid DNA, at a final concentration of 1 mg/mL in saline (0.9% NaCl) formulation. The average yield from 4 different lots was 252.6 ± 26 mg of plasmid DNA per 5 L culture.Final preparations of plasmid DNA were characterized by analytical quality control procedures. Data were consistently reproduced in the 4 lots (data not shown). Expression and biological activity of VEGF121 VEGF121 concentration in supernatants from CHO-K1 cells transfected with four different lots of pVEGF121 ranged between 401 and 683 (541±116) pg/mL of medium. The expressed VEGF121 was shown to be biologically active when assayed in an in vitro model of angiogenesis. Human microvascular endothelial cells grown in the presence of supernatant from pAEC-D2-transfected cells exhibited a small round shape and did not spread. In contrast, treatment with VEGF121-containing supernatants resulted in drastic morphological changes. The cells became elongated, forming thin cords of interconnecting cells (Figure 1). Similar effects were observed with VEGF165 protein. These data demonstrate that VEGF121 is able to mediate dramatic cell reorganization, which would be necessary in vivo for endothelial cell sprouting and tube formation. Angiogenic stimulation in vivo pVEGF121 was assayed for in vivo angiogenesis in a dog hindlimb ischemia model. Animals were subjected to a surgical procedure in order to induce unilateral ischemia. An interval of 10 days was permitted for spontaneous development of collateral vessels. Then, animals received intramuscular injections of pVEGF121 directly in the ischemic muscles. Anatomic and physiological effects of VEGF121 gene transfer were evaluated at different time points up to day 40 (30 days after transfection). Angiography. Representative angiograms recorded from both control and pVEGF121-treated animals are shown in Figure 2. In control animals, collateral artery development in the medial thigh appeared unchanged in serial angiograms recorded at days 10 and 40 (Figure 2, A and B). In contrast, in the pVEGF121-treated group, improvement of collateral artery development was typically observed in angiograms at day 40 when compared with the baseline (day 10) angiogram (Figure 2, C and D). Calf blood pressure ratio. At day 10 after induction of ischemia (immediately before gene transfer), calf blood pressure ratio was similar in both groups (pVEGF121=0.52±0.09, control=0.54±0.13; P=NS). By day 25, a slight improvement in blood pressure ratio was observed in both groups of animals, but differences between groups were not significant (pVEGF121=0.67±0.09 versus control=0.60±0.13; P=NS). However, at day 40, the blood pressure ratio for animals in the pVEGF121-treated group was significantly higher than that for animals in the control group (pVEGF121=0.79±0.05 versus control=0.56±0.14; P<0.01) (Figure 3). Moreover, when compared with baseline, a significant increase in calf blood pressure ratio was found at day 40 in pVEGF121 group (P<0.01), but not in the control group. Vasomotor reserve. Animal responses to an intravenous infusion of nitroglycerin are shown in Figure 4. At day 10 postoperation no animal showed response. At day 25, 50% of control animals partially responded, while 66.7% of pVEGF121-treated animals were partial or total responders. By day 40, responder animals remained 50% for the control group. In contrast, 100% of pVEGF121-treated animals showed partial (33.3%) or total (66.7%) responses. Statistical analysis showed no significant differences (P>0.05) between groups at both, day 25 and day 40. Nevertheless, the biological significance of these results is discussed below. Hematological assays. An important set of hematological variables were measured in order to evaluate possible side effects of pVEGF121 on animal homeostasis (Table 1). The statistical analysis showed not significant differences between groups for all variables at all measurement points. In addition, no statistical differences were found at days 20 and 40 with respect to day 10 (baseline). Moreover, all variables remained within or close to the physiological ranges (Tvedten, 1981).
Discussion The present study reports on the construction of a plasmid DNA expressing the 121-amino acid isoform of VEGF and the successful gene transfer and expression of pVEGF121 in an animal model of hindlimb ischemia, with evidence of increased collateral vessel development, and improvement in hemodynamic and physiological deficits induced by ischemia. pVEGF121 plasmid was constructed as a derivative of pAEC-D2 (Herrera et al. 2000), and purified from bacterial culture of E. coli DH10B strain. The expression of the transgene was achieved by transfection of CHO-K1 cells and verified by anti-VEGF ELISA. The biologic activity of the expressed VEGF121 was confirmed by stimulating human microvascular endothelial cells with conditioned media from pVEGF121 transfected cells in an in vitro model of angiogenesis. Angiogenic effects of pVEGF121 were evaluated in vivo using a dog model of hindlimb ischemia. A similar model has been described in rabbits, including reduced TcO2 (Baffour et al. 1992), increased femoral venous lactate (Pu et al. 1994), and skeletal muscle necrosis (Baffour et al. 1992). However, our model exhibits several advantages when compared with the rabbit one. In the canine model, the vessels are more accessible for being dissected free. The anatomic system from the iliac sector and below includes collateral arteries, such as the internal saphenous, allowing the setting of a more severe experimental ischemia (Rakue et al. 1998). Regarding anesthetics and surgical procedures, dogs are more resistant than rabbits. In fact, no animal died during our study in spite of the strong bleeding imposed by the procedure. Moreover, angiography can be performed by direct puncture after palpation of the aorta beat, and no last generation equipment is required. In addition, gene transfer efficiency is usually inversely related to host size because of limited tissue diffusion of the gene transfer vectors and larger volumes of the transfected tissues (Yla-Herttuala and Alitalo, 2003). In this regard, the dog model is expected to have a higher predictive value because it is closer to the human size. In our study, the ischemic condition was confirmed 10 days after the surgical procedure by measuring the calf blood pressure ratio. When compared to intact limbs, operated ones showed a mean decrease of 53% in systolic pressure (data not shown). pVEGF121 was intramuscularly injected into the ischemic limbs. Augmented collateral circulation was documented by angiographic analysis 30 days after gene transfer (day 40). Improvements in physiological parameters regarding macro- and microcirculation were evidenced by analyses of the calf blood pressure ratio and the vasomotor reserve, respectively. The circulatory system is composed of at least two basic components in the arterial segment; macrocirculation, including large and medium diameter vessels; and microcirculation, concerning small vessels and capillars. Macrocirculation is related to conducting and compensatory functions; while nutrient interchange is the main function of microcirculation. Both of them are necessary and complement each other to achieve adequate blood circulation and tissue nutrition (Norgren, 2000). Analysis of calf blood pressure ratio at day 40 documented a statistically significant increase in the pVEGF121-treated group, when compared to the control group, indicating an improvement in blood flow at large and medium diameter vessels. The pVEGF121-treated group showed an increase in pressure ratio of 0.27 at day 40 with respect to baseline (Fig. 3), being equivalent to a 20 mm Hg increase in the limb systolic pressure. This is a remarkable improvement of high clinical value. Vasomotor reserve assay suggested amelioration in blood availability at the microcirculation level in pVEGF121-treated animals. Despite the lack of statistical significance, the biological relevance of these results should be considered. The fact that all pVEGF121-treated animals, as opposed to 50% in the control group, were responders to the nitroglycerin infusion indicates a tendency for a positive effect of pVEGF121. Furthermore, responses in the pVEGF121 group were 33.3% partial and 66.7% total (2 and 4 animals, respectively). However, in the control group, 2/3 of responder animals reacted partially and only 1 animal showed total response (Fig. 4). This finding could have important implications when considering establishing a therapeutic approach for critical limb ischemia. Increasing the number of animals in the study is recommended in order to achieve statistical significance. Improvements observed in the control group regarding both, calf blood pressure ratio and vasomotor reserve, are probably due to ischemia-induced endogenous angiogenesis (Semenza, 2001). Our present results are in agreement with previous studies that have successfully applied intramuscular gene transfer of VEGF165 in the rabbit hindlimb ischemia model (Tsurumi et al. 1996; Tsurumi et al. 1997). Percutaneous arterial gene transfer by means of a hydrogel-coated balloon catheter (Takeshita et al. 1996b; Takeshita et al. 1996c) has also been reported for therapeutic angiogenesis. Despite their encouraging findings, arterial gene transfer has certain limitations for treating lower limb ischemia. In these patients, several factors such as extensive ischemia, atherosclerosis, and extensive calcified deposits, may compromise the access to a satisfactory arterial donor site in the lower extremity circulation. Even if arterial access is possible, it is often limited to the uppermost portion of the limb, ³60 cm from sites in the distal limb at which ischemia is most profound. In contrast, intramuscular gene transfer represents a less invasive alternative, positioning the putative sites of VEGF synthesis in closer proximity to the ischemic focus. The ready access to ischemic skeletal muscle, make this approach suitable for the treatment of critical limb ischemia. Striated muscle has been shown to take up and express foreign genes transferred in the form of naked plasmid DNA. Upon intramuscular injection, such plasmids remain in a nonreplicative, unintegrated, circular form (Danko and Wolff, 1994), so this strategy is unlikely to be complicated by insertional mutagenesis. The promising results of preclinical experiments using intramuscular gene transfer of VEGF165 (Tsurumi et al. 1996; Tsurumi et al. 1997) have been confirmed by early clinical trials (Baumgartner et al. 1998; Isner et al. 1998). The limiting factor associated with the use of naked DNA for direct intramuscular gene transfer has been the level of gene expression required for phenotypic modulation (Ledley, 1995). However, it has been shown that VEGF121 and VEGF165 are efficiently synthesized (up to 3 ng/mL medium) in spite of low transfection efficiency (5%) of VEGF-carrying plasmids in vascular smooth muscle cells (Dulak et al. 2000). Additionally, the secreted nature of VEGF is another factor contributing to the success of intramuscular gene transfer with naked DNA. A previous work with reporter genes (Losordo et al. 1994) has indicated that low efficiency of transfection with genes encoding for secreted proteins may achieve biologically meaningful effects not achieved by transfection with genes encoding for intracellular proteins. Thus, even if VEGF gene transfer is limited to a relatively small number of cells, the expression levels and the paracrine effects of the secreted gene product may be sufficient to achieve a meaningful biological effect. The ischemic environment of the transfected muscle is another factor contributing to transgene uptake. Previous studies (Takeshita et al. 1996a; Tsurumi et al. 1997) have shown that transfection efficiency of intramuscular gene transfer increases more than fivefold when the injected muscle is ischemic. On the other hand, paracrine induction of KDR in endothelial cells exposed to media conditioned by hypoxic myoblasts, has been demonstrated (Brogi et al. 1996). This finding acts to amplify the impact of any given concentration of VEGF. Thus, ischemia further contributes to the success of therapeutic angiogenesis, independent of transgene uptake, by modulating VEGF receptor expression. From a safety standpoint, a localized and transient gene expression is desirable, in order to avoid pathologic angiogenesis at sites different from the site of gene transfer. VEGF has been implicated in the pathogenesis of proliferative retinopathy of diabetic patients (Boulton et al. 1998) and in the growth and development of latent tumors (Siemeister et al. 1998). The site-specific nature of the angiogenic response following systemic administration of recombinant VEGF has been documented (Bauters et al. 1995). Previous studies with VEGF165 have shown that the transgene was expressed in the transfected muscles for as long as 14 days after intramuscular gene transfer, as assessed by RT-PCR (Tsurumi et al. 1996; Tsurumi et al. 1997). Once obtained a satisfactory clinical benefit, early cessation of gene expression assures that the recipient is not exposed indefinitely to increased levels of VEGF. In the present study, no modifications were detected in several hematological variables as a result of VEGF121 gene transfer. This finding represents a safety feature of VEGF gene therapy, in that no side effects could be associated to the administration of pVEGF121 plasmid. A recent clinical trial has reported that adenovirus-mediated gene transfer of VEGF121 improves lower limb flow reserve and endothelial function in patients with peripheral arterial disease (Rajagopalan et al. 2001). Other authors demonstrated that adenovirus-mediated VEGF121 gene transfer stimulates angiogenesis in normoperfused skeletal muscle, and the newly formed vessels preserve tissue perfusion after induction of ischemia (Gowdak et al. 2000). The same approach has been used in several studies with VEGF121 to promote angiogenesis in the ischemic myocardium of animals (Patel et al. 1999; Lee et al. 2000), and patients (Rosengart et al. 1999a; Rosengart et al. 1999b). However, the administration of large amounts of virus can lead to massive immune responses that could cause serious immunopathology. Even when the virus is administered in relatively small amounts, the foreign proteins presented by the virus can induce immune responses that could decrease subsequent sensitivity to the beneficial effects of the transgene delivered by the virus if repeatedly administered, or could possibly lead to immune-mediated tissue damage (Epstein et al. 2001). Unlike viral vectors, no evidence of an immune response has been recognized in animal experiments using naked plasmid DNA (Nabel et al. 1993). DNA is subsequently denatured and is likely to be removed by the reticuloendothelial system. Previous studies suggest that histones may account for a significant proportion of the antigenicity associated with DNA (Burlingame et al. 1994). In the case of plasmids generated from E. coli, however, histones are absent from the recombinant DNA, thus further reducing the likelihood of an immunologic response. Most of the previous works on VEGF gene transfer using naked plasmid DNA have used the 165-amino acid isoform (Tsurumi et al. 1996; Tsurumi et al. 1997; Baumgartner et al. 1998; Isner et al. 1998). Meanwhile, VEGF121 gene transfer has been performed by means of adenovirus (Rosengart et al. 1999a; Lee et al. 2000; Rajagopalan et al. 2001). This is the first study reporting on intramuscular gene transfer of the 121-amino acid isoform of VEGF as naked plasmid DNA in a dog model of hindlimb ischemia, taking into account the higher robustness and predictive value of this model (Rakue et al. 1998). VEGF121 and VEGF165 are the best studied isoforms in therapeutic angiogenesis research. These isoforms markedly differ in their secretion pattern. VEGF121 does not bind to heparin and is freely soluble in the conditioned medium of transfected cells; while VEGF165 shows heparin binding ability and, although diffusible after secretion, a significant portion remains bound to the cell surface or extracellular matrix (Park et al. 1993). It has been speculated that angiogenesis induced by VEGF may be differentially dependent upon the extent to which each particular isoform is freely secreted and soluble. However, biologic equivalence among 121, 165, and 189 VEGF isoforms has been demonstrated for in vivo angiogenesis (Takeshita et al. 1996b). This fact may be interpreted to support previous observations that plasmin can cleave VEGF165 and VEGF189 and release active soluble proteolytic fragments having a size similar to or smaller than the intact 121 isoform (Houck et al. 1992; Plouet et al. 1997). The proteolytic cascade of plasminogen activation plays a key role in angiogenesis (Pepper, 2001); it is, therefore, possible for it to cleave the longer forms of VEGF and release soluble VEGF121-like species, being the final common mediator of angiogenesis in vivo. The present research provides evidences that direct intramuscular injection of naked plasmid DNA encoding VEGF121 is an effective method for gene transfer in an animal model of hindlimb ischemia, resulting in increased collateral vessel development and tissue perfusion. Thus, our results suggest the feasibility of pVEGF121 plasmid to promote therapeutic angiogenesis in patients with critical limb ischemia.
Note: Electronic Journal of Biotechnology is not responsible if on-line references cited on manuscripts are not available any more after the date of publication. Supported by UNESCO / MIRCEN network © 2003 by Pontificia Universidad Católica de Valparaíso -- Chile The following images related to this document are available:Photo images[ej03025f1.jpg] [ej03025f3.jpg] [ej03025f4.jpg] [ej03025f2.jpg] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}