 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
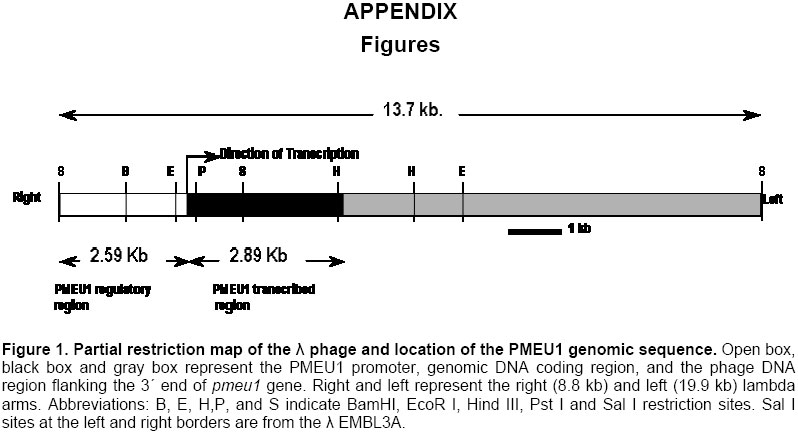
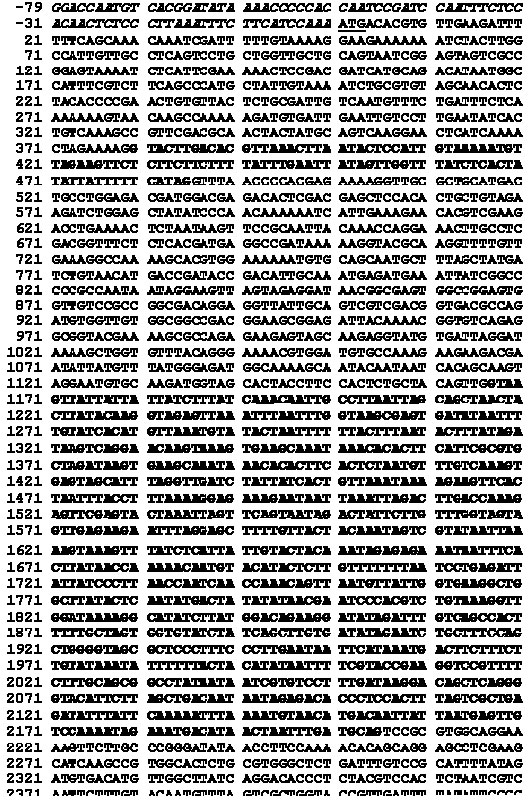
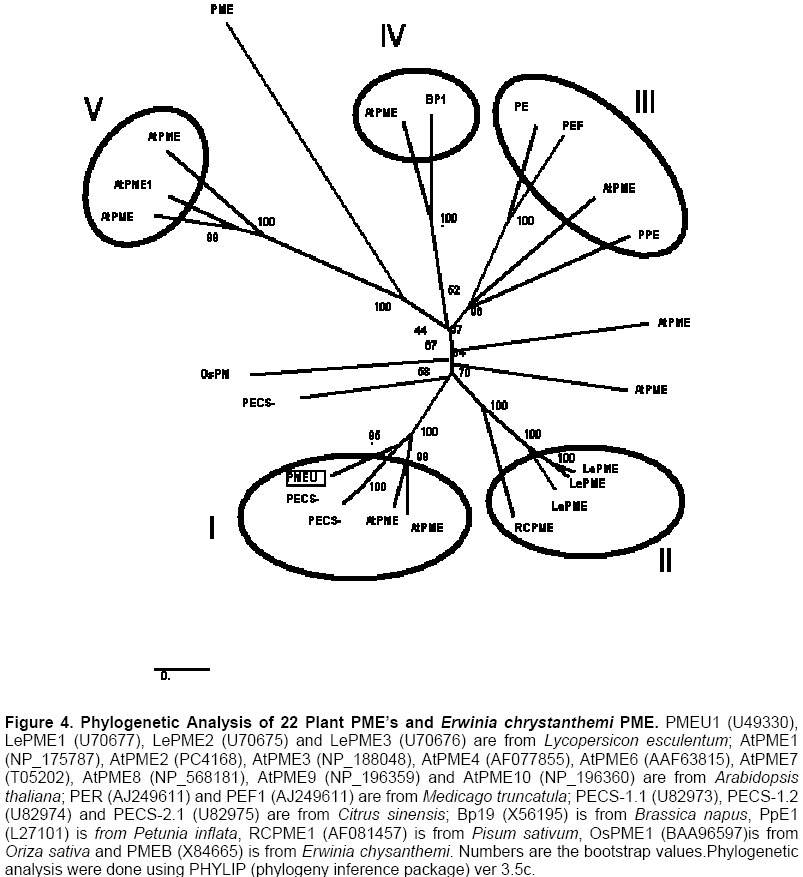
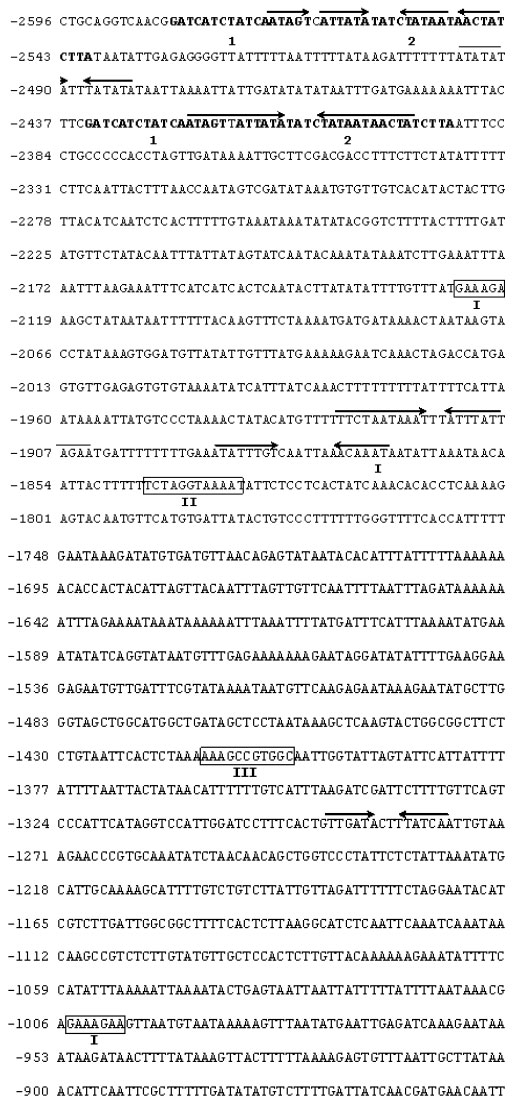
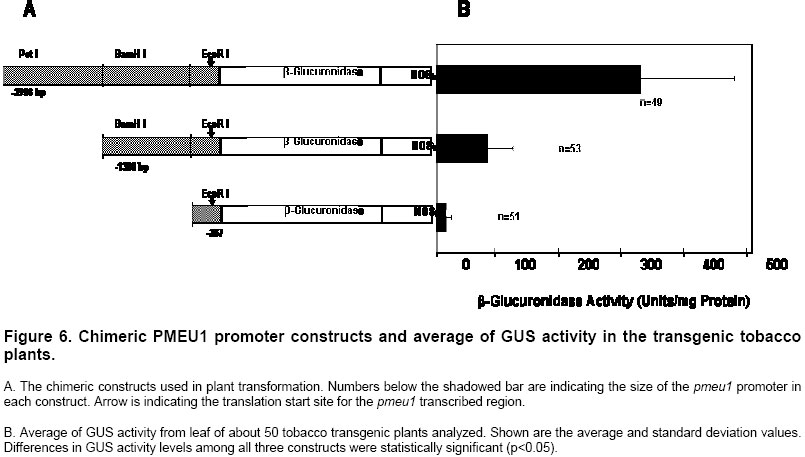
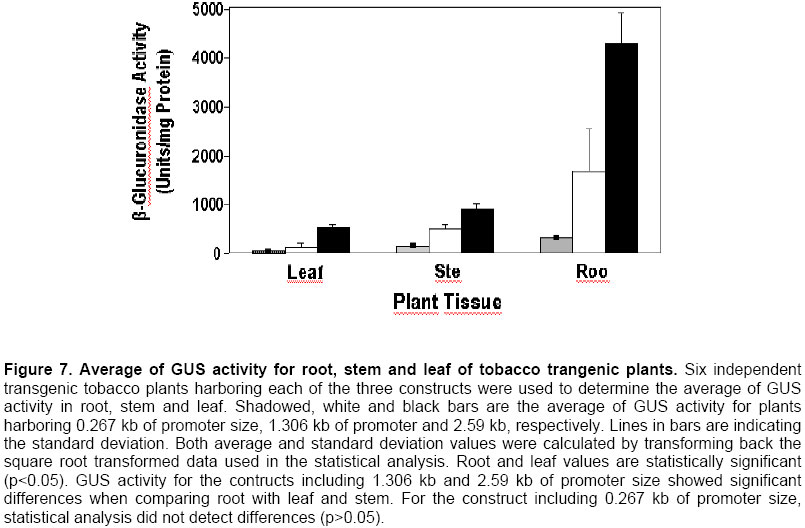
Electronic Journal of Biotechnology, Vol. 7 No. 1, April 15, 2004 Isolation and study of a ubiquitously expressed tomato pectin methylesterase regulatory region Martín-Ernesto Tiznado-Hernández*1, Joel Gaffe2, Avtar K. Handa3 1Departamento
de Tecnología de Alimentos de Origen Vegetal,
Centro de Investigación en Alimentación y Desarrollo, A.C.,
Carretera a la Victoria km. 0.6,
Apartado Postal 1735,
Hermosillo, Sonora, 83000, México,
Tel: 52 662 80 00 55,
Fax: 52 662 280 04 22,
E-mail: tiznado@cascabel.ciad.mx Received September 28, 2003 / Code Number: ej04003 Abstract Pectin methylesterase (PME) is an enzyme located in the plant cell wall of higher plants whose physiological role is largely unknown. We had isolated a PME gene from a tomato genomic library, including 2.59 kb of 5´ flanking region and the coding region. Both coding and promoter region were sequenced and computer analyzed. Tobacco transgenic plants were created harboring constructs in which 2.596 Kb, 1.306 Kb and 0.267 Kb sizes of the promoter were driving the expression of β-Glucuronidase gene (GUS). GUS activity was studied by histochemical and fluorometric assays. Two introns of 106 and 1039 bp were found in the coding region and phylogenetic analysis placed this PME gene closer to genes from Citrus sinensis and Arabidopsis thaliana than tomato fruit-specific PME genes. In the promoter, it was found direct repeats, perfect inverted repeats and light responsive elements. GUS histochemical analysis showed activity in all plant tissues with the exception of pollen. The reduction in the promoter size induced a reduction in GUS activity in root, stem and leaf. Furthermore, root and leaf showed the highest and lowest activity, respectively. We had isolated a tomato PME gene with novel characteristics as compared with other known PME genes from tomato. Keywords: pectin Methylesterase, promoter analysis, tobacco transgenic plants, tomato. Pectin methylesterase (PME) is an enzyme that have been found in every plant tissue analyzed (Lineweaver and Jansen, 1951; Rexova-Benkova and Markovic, 1976), several fungi (Christgau, et al. 1996; Mendgen, et al. 1996), bacteria (Plastow, 1988; Barras et al. 1994) and even insects (Ma et al. 1990; Shen et al. 1999). In higher plants, it is known to be a cell wall associated protein and several of the PME cDNA available in the literature, are known to have toward the N-terminal sequence, a characteristic signal peptide which is thought to help in targeting the protein to the plant cell wall (Gaffe et al. 1997 and references therein). PME catalyzes the deesterification of galactosyluronate methylesters of pectins, releasing protons and methanol into the media (Frenkel et al. 1998). Despite the biochemical mode of action of PME is well known, it have been difficult to demonstrate any role for PME in the physiology of plants. However, several hypothesis had been proposed: pollen germination and/or tube growth (Mu et al. 1994), abscission (Sexton and Roberts, 1982), regulation of cell enlargement through changes in the plant cell wall Donnan potential (Ricard and Noat, 1986), fruit softening during postharvest fruit ripening (Zeng et al. 1996) and plant defense (Chamberland et al. 1991; Wietholter et al. 2003). Furthermore, strong experimental evidences had been provided to suggest that a PME role in the release of cells from the root cap (Stephenson and Hawes, 1994; Wen et al. 1999), plant pathogenesis (Collmer and Keen, 1986; Mendgen, et al. 1996; Nun et al. 1996; Valette-Collet et al. 2003), plant systemic infection by tobacco mosaic virus (Chen and Citovsky, 2003) and maintenance of the tomato fruit tissue integrity during postharvest shelf life (Tieman and Handa, 1994). However, the actual physiological role of PME is still matter of controversy. Four papers to our knowledge had been published in which the cloning of regulatory sequences of PME from higher plants was described. However, in all of them DNA comparison by computer was the only tool used to prove that the gene located downstream was indeed encoding a pectin methylesterase. Albani et al (1991) reported the finding of a genomic clone from Brassica napus which contains the PME gene and its 5´ upstream regulatory region. Studies were conducted using a piece of the gene located dowstream as a probe. This gene was found to be expressed mainly during pollen development. Two putative PME promoter regions were cloned from Brassica campestris (Kim et al. 1997). Study of their sequence found them to have high homology with the previously reported promoter PME from Brasicca napus (Albani et al. 1991). Further, a sequence motif similar to the one known to exist in two tomato pollen-specific promoter was located. Tobacco transgenic plants with constructs containing two different promoter sizes from one of those two promoter available were made. Expression of the GUS gene was only detected during developing and mature pollen grains germinated in vitro. Recently, the cloning of two 5´ upstream region of PME from Citrus sinensis was published. Northern blot analysis showed that both DNA regulatory region are active in most of vegetative tissues (Nairn et al. 1998). In our laboratory, we have cloned a 13.7 kb. genomic DNA from tomato containing a 2.59 kb. of DNA 5´ flanking region, along with all the PME genomic clone. Identification of the protein encoded by the gene downstream was made by creating tobacco transgenic plants over expressing the PME cDNA (Gaffe et al. 1997) and comparing the sequences of the genomic and cDNA regions. In this work, we describe the study of a 5´ flanking region of a PME gene called pmeu1 (which stands for PME ubiquitous one) using computer tools and tobacco transgenic plants. Materials and Methods Cloning of the genomic fragment The plaque lift technique was used to screen 810,000 clones of a tomato cv ‘Cherry’ genomic library made in EMBL3A λ phage with a radiolabeled small piece of PME; cloned by RT-PCR from tomato roots poly-A+ mRNA (Gaffe et al. 1995). We found several hybridizing, putatively positives, plaques. From every plate, we made two lifts and only that plaques producing signal in both lifts were chose to continue. After four rounds of purification and screening, one plaque turns out to be positive. Elimination of the bacteria present in the agar was made by using chloroform-containing SM buffer. For phage amplification, E. coli strain LE 392 was infected with the phage after cultured in LB media. DNA isolation from the phage was made through a phenol chloroform protocol (Ausubel et al. 1988). Digested DNA with several restriction enzyme was separated by electrophoresis and blotted into nylon membranes. These DNA blots were probed with the PME cDNA complete sequence available to locate the phage DNA region encoding the genomic PME gene and the region 5´ upstream. Sal I and EcoR I digested DNA fragments were subcloned into pBSKS (+/-) vector (STRATAGENE CLONING SYSTEMS. La Jolla, CA). The EMBL3A λ phage library screened was created using the Sal I restriction site. Because of this, digested DNA fragments using Sal I were used to calculate the size of the tomato DNA inserted into the phage isolated, found to be 13.7 kb. All the procedure above mentioned was performed essentially as described (Sambrook et al. 1989) unless otherwise indicated. DNA sequencing of the promoter and the genomic coding region Nested unidirectional deletions of the 5´ upstream DNA Sal I fragment were made by following the recommendations of the company (Erase-a-Base® System, Promega Corporation, Madison, WI). Deleted clones with about 250 bp of size difference were used for DNA sequencing using the T3 universal primer by the Sanger dideoxy chain termination technique following the recommendations (Sequenase Kit, United States Biochemical Corporation, Cleveland, Ohio). Second strand sequencing was determined by the DNA sequencing facility of IOWA State University by using primers designed at proper positions in the sequence (Iowa State University, Ames, Iowa). Three chimeric constructs driving the β-glucuronidase gene (uidA) under different sizes (2.596 Kb, 1.306 Kb and 0.267 Kb) of the promoter region were created by transcription fusion through the insertion of two stop codons in between the ATG of the pmeu1 gene and the ATG of the uidA gene. Every chimeric construct was ligated into the promoterless binary vector pBI101.3 (Bevan, 1984). This plasmid includes the neomycin phosphotransferase gene (NPTII) which confers resistance to kanamycin to be used as selectable marker. Furthermore, in this plasmid the DNA introduced is located between the right and left borders of the T-DNA, which allows the transference into the plant genome by Agrobacterium infection (Hooykaas, 1989; Zupan and Zambryski, 1995; Nester et al. 1996). Proper insertion of the different promoter sequences into the plasmid was confirmed by DNA digestion using suitable restriction enzymes and PCR using primers designed against sequences in the PME promoter and uidA gene. Every chimeric construct created included 150 bp in between the ATG of the pmeu1 gene and the ATG of the uidA gene, containing sequences from the pmeu1 gene and pBSKS(+/-) phagemid and pBI101.3 binary vector. Sequencing between the two ATG’s was used to verify the presence of two stop codons and to corroborate the transcription fusion of the two ATG. Mobilization of the pBI101.3 plasmid into Agrobacterium LBA4404 was performed by triparental mating using the broad-host helper plasmid pRK2013 (Ditta, 1981). Agrobacterium transconjugants were screened on plates containing a mixture of kanamycin and rifampicin antibiotics in YEP media (Sambrook et al. 1989). Verification of the mobilization of the constructs was made by purification DNA from Agrobacterium following the recomendations (Wizard Minipreps, Promega Corporation, Madison, WI) and digestion with proper restriction enzymes. Tobacco (Nicotiana tabaccum W38) young leaves and tomato (Lycopersicon esculentum cv Ohio 824) cotyledons were infected with Agrobacterium by using the leaf disk technique (Mathis and Hichee, 1994) and selection for transformants was done by using kanamycin in the media. After induction of roots, about 50 primary independent transformant plants growing in vitro harboring every of the three constructs were selected at random to measure GUS activity in leaf. This was done using the fluorometric technique (Jefferson et al. 1987) with a Perkin Elmer LS5 fluorometer. Quantification of reaction product was done by using a 4-methylumbelliferone standard curve. Also, six independent transgenic plants were used to measure GUS in root, stem and leaf tissues. Every GUS measurement was done at least three times. For enzymatic specific activity, protein determination was made using Bradford (1976) with bovine serum albumin as standard. In order to examinate the GUS presence in different tissues, at least 20 primary transgenic plants harboring the different constructs, were vacuum infiltrated with a 1.9 µM solution of 5-Bromo-4-Chloro-3-Indolyl-Glucuronide (Jefferson et al. 1987) as described (Mandel et al. 1995). Tobacco flowers in anthesis were collected from plants growing in the greenhouse and transported immediately to the laboratory. Anthers were cut and only that pollen released by a gently shaking was used for germination studies. Pollen was germinated using the Brewbaker and Kwack solution as described (Brewbaker and Kwack, 1963). Histochemical GUS staining was performed after four hours of pollen germination. Germination solution was changed by the GUS staining solution and left at 37ºC for at least 18 hrs before examination for GUS staining. DNA sequence from the different pmeu1 5´ flanking region deletions were joined together using DNAsis (Hitachi Software Engineering Co., LTD., 1991). Comparison between the pmeu1 cDNA and pmeu1 genomic clone was performed using Harr plot analysis with DNAsis software. Presence of known cis-acting elements was determined using the programs MathInspector ver. 2.2 (Quandt et al. 1995), TFSEARCH ver. 1.3 (Parallel Application, Tsukuba Laboratory, RWCP, Japan), Signal Scan ver. 4.05 (Prestridge, 1991) and Pattern Search (Wingender et al. 1996; Wingender et al. 1997). Percent of identity among the different PME promoters and PME transcribed regions were determined using Align (Myers and Miller, 1988). Alignment of deduced amino acid sequences was performed using GCG’s Pileup Program (Genetics Computer Group, Madison, WI). Multiple sequence alignment was performed using CLUSTAL W (Thompson et al. 1994). DNA direct repeats for the tomato PMEU1 promoter were determined using Proscan ver 1.7 and repeats from GCG software ver. 9.0 (Genetics Computer Group, 1995). Perfect inverted repeats (mirror repeats) were located using Palindrome from GCG software ver 9.0 (Genetics Computer Group, 1995). Putative TATA box was located by Signal Scan ver. 4.05. Phylogenetic analysis were done using the phylogeny inference package (Felsenstein, 1989; Felsenstein, 1993). Comparison of leaf GUS activities for the three constructs and for the different tissues was made by variance analysis using a completely randomized design for unbalanced number of repetitions. Tukey test was used when needed to find differences among means. Because it is known that the GUS enzymatic activity in populations of first-generation transgenic plants does not follow a normal distribution (Nap et al. 1993), we performed a Box-Cox transformation before variance analysis. From here, we learned that a square root was a suitable transformation to bring the GUS activity parameter into normality. Statistics reported in this paper represents the back transformation of the square root transformed data. All statistical analysis were performed using the SAS software (SAS Institute Inc. Cary, N.C.). Results Isolation and characterization of PMEU1 gene The cloning and characterization of the entire PMEU1 tomato cDNA has been previously reported (Gaffe et al. 1996; Gaffe et al. 1997). The next step lead us to the isolation and characterization of the genomic fragment containing the PMEU1 gene. An EMBL3A phage of a tomato genomic library (VNTF cherry) was screened using 300 bp cDNA fragment corresponding to the conserved PME domain in PMEU1 (Gaffe et al. 1996; Gaffe et al. 1997). Four rounds of phage amplification allowed us to purify a single positive clone. Subcloning, analysis by restriction mapping and DNA blot of the tomato genomic DNA fragment contained in the EMBL3A phage indicated that the size of the inserted tomato genomic DNA is 13.7 kb and the PMEU1 gene was found to be located toward the 5´ region, spanning 5.28 kb. In Figure 1, it is presented the organization of the EMBL3A clone containing the PMEU1 gene This region includes 2.59 kb of DNA regulatory region and 2.89 kb of DNA transcribed region, shown as white and black areas. In the figure it is also shown the location of the right and left lambda phage arms and the main restriction sites. DNA sequence of the transcribed region of PMEU1 gene In Figure 2, it is shown the sequence of the PMEU1 genomic clone (GenBank Accession Number: AY046596). In italics, it is presented the 5´ untranslated region (Gaffe et al. 1997) and the partial 3´ untranslated region. In bold, it is shown the sequence of the two introns present. Underlined, it is presented the translation start site and stop codon (TAA). Double underlined it is shown the putative polyadenylation signal and polyadenylation site (GT). The polyadenylation signal was found to follow the plant consensus sequence AAUAAA (Li and Hunt, 1995). The two introns present are of 106 and 1039 bp in length. Both of them showed a significantly higher composition of U’s with respect to the flanking exon sequences. This is a characteristic known to be present in many plant genes (Ko et al. 1998). Intron-exon lrganization of PMEU1 and other PME genomic clones The intron-exon structure of the PME genomic sequences available has been analyzed. The splice junction of all the clones conform to the GT/AG boundary rule for the 5´ donor and 3´ acceptor site (Liu and Filipowicz, 1996). The intron size range from 72 to 1577 bp and the exon from 117 to 1353 bp. The average value for intron and exon size is 109 and 519, respectively. Seventeen clones have only one or two introns. Three putatives PME genomic sequences from Arabidopsis contains four introns and show a level of similarity with PMEU1 of around 50%. Further, AtPME7 with five introns is more closely related to PMEU1 (64.9% of similarity). These observations suggest that there is not a simple relationship between the phylogenic distance and intron number in the different PME genomic clones. The position of one intron, relative to the deduced amino acid sequence, is conserved in 19 out of the 22 plant PME genomic sequences. This intron is located 17 amino acid residues upstream of the PME signature sequence GPXKHQAVALR; observed in the rice genomic clone as well (Figure 3 a, b, c, d). This observation suggest that monocots and dicots share a common ancestor. The other three clones (AtPME8, AtPME9 and AtPME10) are clustered together in one group by the phylogenic analysis (Figure 4) which agrees with the lack of the intron located at the same distance from the signature sequence and the common characteristic of the presence of four introns. Phylogeny analysis among PMEU1 and other plant PME genes Deduced amino acid sequences of 22 plant PME genes as well as PMEB from Erwinia chrysanthemi were included in our study. The plant PME genes were chosen based in published data providing experimental evidences or presence of the full genomic sequence from the Arabidopsis thaliana genome project from which some of the PME genes were included. One of the pectin methylesterase genes from Oriza sativa was included to be able to compare with a PME from monocots. Furthermore, the gene from E. chrysanthemi was chosen in order to compare PME from plants with a distantly related PME and also to have a control in the phylogenetic analysis. The PMEU1 gene includes 2900 bp and a theoretically deduced open reading frame of 583 amino acids (Figure 3 a, b, c, d). Several sequences shorter than 400 bp like PECS-1-2 from Citrus sinensis, are known to be partial. However, PPE1 sequence from Petunia inflata is shorter than 400 bp and still encodes a full polypeptide. Sequence alignment of these different encoded polypeptides indicate that the N-terminal half of these clones is loosely conserved compared with the C-terminal half, involved perhaps in the PME catalytic activity (Figure 3 a, b, c, d). Because of this, a final alignment, edited to represent only the phylogenetically relevant fraction of the sequences was used to derived a phylogenetic tree (Figure 4). Based on this phylogenetic analysis, we organized up to 18 genomic clones in five groups. Five PME genomic clones from various origins can not be associated with any of these groups. The lack of association of PME from Erwinia chrysanthemi with other plant PME’s was something expected, however, it is interesting that the clone PECS-2.1 from Citrus sinensis is distantly related with the two clones PECS-1.1 and PECS-1.2 from the same source that clustered together with the PMEU1 clone. This phylogenetic analysis indicates that PMEU1 belong to a group containing two Citrus sinensis PME genes, PECS-1.1 and PECS-1.2 and two Arabidopsis thaliana genes, AtPME2 and AtPME3; however, it is distant from the three tomate PME genes expressed only in tomato fruit tissues: LePME1, LePME2 and LePME3 (Harriman et al. 1992), suggesting the PMEU1 is a gene evolved to have a different and novel function. However, due to the limited amount of information concerning the expression of these genes, we can not establish a clear relationship between these groups of PME genes and their possible function. In Figure 5 (a, b) it is shown the 2.59 kb. PMEU1 promoter sequence (GenBank Accession Number AY050764). Computer study of this sequence showed several features commonly present in DNA regulatory sequences. The largest direct repeats within the promoter sequence, are shown underlined and numbered. Mirror repeats are shown with arrows in opposite directions. Putative cis-acting elements are shown boxed and roman numbered. The putative TATA box is shown double underlined. In bold, it is shown the transcription start site. Study of the 5´ region of this sequence did not indicate the presence of elements commonly present in the 3´ region of genes, suggesting that the PMEU1 promoter region could be larger than 2.59 kb. The number of direct repeats located by computer in the PMEU1 promoter varied with the size of the fragment, in such a way that it was found only one for repeats consisting of 17 and 26 bp, four for repeats with 12 bp, three for repeats with 11 bp and greater than 1000 for repeats with 5 bp (data not shown). However, the significance of this repeats within the PMEU1 promoter remains to be elucidated. We also locate in the promoter sequence several perfect inverted repeats o mirror repeats, depicted in Figure 5 (a, b) as arrows pointing in opposite directions. It is interesting that the longest inverted repeats is contained within the longest direct repeats. As in the case of the direct repeats, the function of these inverted repeats, if any, is unknown. Short sequences with resemblance to known cis-acting elements present in other ADN regulatory regions were located in the PMEU1 promoter sequence. In Figure 6 are included only the ones with the highest degree of similarity. Two copies of the sequence GAAAGA shown to confer responsiveness to red light in the phytochrome A3 promoter (Bruce et al. 1991) are present in PMEU1 promoter (box I). Also, one copy similar to the sequence GTGAGGTAATAT, known to be regulated by light (Fluhr and Dankekar, 1986; Green et al. 1987) was found (box II). Furthermore, we found regions similar to a G-box (box III), shown to be light inducible (Schindler et al. 1992). Also, it was located a sequence similar to an abscisic acid responsive element (box IV) (Guiltinan et al. 1990). As can be seen from above, three of the four putative cis-acting elements located are known to be regulated by light. Experiments to show whether PMEU1 promoter is regulated by light deserves further attention. However, still the function of this cis-acting elements within the PMEU1 promoter is largely theoretical and experimental evidences to confirm any function of these sequences remains to be provided. We were able to locate a putative TATA box 44 bp upstream of the transcription start site (Figure 5 (a, b)). However, as mentioned for the other elements above described, the confirmation of this region as actual TATA box still need to be experimentally probed. We did not find the presence of a CAAT box, although it had been shown to be present in several promoter of plant genes (Joshi, 1987). Paired comparisons among the DNA sequence of the PMEU1 promoter with sequences of PME promoters from Brassica campestris (GBAN215-6 and GBAN215-12), Brassica napus (Bp 19), Citrus sinensis (CsPME1 and CsPME3) and Arabidopsis thaliana (AtPME1) did not showed any special pattern or similarity with any of the promoters included in the analysis. Indeed, all the pair comparisons showed around 50% of identity. Further, analysis by multiple sequences alignment among all PME promoters failed to locate an homologous region in common to all of them (data not shown). With the goal to test whether the 2.59 kb. DNA region located in the 5´ flanking region of the PMEU1 genomic coding region represent an active promoter, we created several tobacco transgenic plants expressing chimeric constructs in which 2.59, 1.3 and 0.267 kb of promoter sequence is driving the expression of the reporter gene uidA encoding the β-glucuronidase enzime. In Figure 6 it is shown the three constructs made along with the average of leaf GUS activity for about 50 independently tobacco transformed plants growing in vitro and expressing the corresponding construct. From the graph, it is clear the trend: the bigger the piece of the promoter, the higher the activity of the uidA gene. Statistical analysis of root squared-transformed data found differences among all of them (p<0.05). Histochemical staining of many independent primary tobacco transgenic seedlings showed activity in leaf, stem and roots of the plants. We also found activity in petals and sepals. However, no activity was detected in pollen grain or in vitro germinated pollen (data not shown). In Figure 7, it is shown the average values of GUS activity for root, stem and leaf of six independent tobacco plants harboring every of the three constructs. The effect of reducing the size in the PMEU1 promoter for the different tissues analyzed followed the pattern already observed in leaf. The decrease in the size of the PMEU1 promoter region reduce its transcriptional activity in all differentiated tissue analyzed. Statistical analysis found significant differences (p<0.05) among the root tissues from plants harboring the different sizes of the promoter. For stem tissues, significant differences were found only between plants with 0.267 kb and 2.59 kb of promoter size. This result is most likely due to the few independent transformants used in the analysis. However, the trend is clear and similar in all plant differentiated tissues analyzed. Discussion We have cloned and analyzed a genomic DNA region containing an almost complete and novel PME gene. Several tools were used to probe that this region encodes the genomic sequence of a PME gene. Comparison of the sequences of PMEU1 genomic coding region with the PMEU1 cDNA already cloned showed that both are identical with the exception of the intron sequences located in the genomic clone. Further, analysis of the cDNA sequence using BLAST resulted in high similarity with several DNA regions encoding PME genes. Also, transgenic plant overexpressing the PMEU1 cDNA under the control of the cauliflower mosaic virus showed higher levels of PME activity as compared with control plants. It was also shown that this high level of PME activity correlated with the presence of a band hybridizing with a PMEU1 specific probe (Gaffe et al. 1997). The PMEU1 gene is presented in the tomato genome as a single copy (Gaffe et al. 1997), in contrast with other PME genes published which had been shown to form clusters (Richard et al. 1996; Turner et al. 1996). We perform several experiments to find another copy of the gene, like increasing the number of plaques screened and using probes from the 5´ end of the gene with unsuccessful results. Also, DNA blot analysis of the 8.4 kb of the 3´ end of the DNA inserted in the phage did not show any hybridization with PMEU1 probe even under low stringency conditions (data not shown). Further, DNA blot analysis of the tomato genome using EcoR I as restriction enzyme showed one band hybridizing to a 6.0 kb band, which correspond precisely with the fragment released from the DNA phage and shown to hybridize with the PMEU1 specific probe (data not shown). Taken together, these evidences support that the PMEU1 gene is presented as a single copy in the tomato genome and that it is part of the DNA contained by the isolated phage from the genomic library. Comparison of the PMEU1 genomic coding region with the PMEU1 cDNA sequence showed the presence of two introns with 106 and 1039 bp in size (Figure 2). We compared the structure of genomic regions encoding PME genes in regard to the number and size of introns. The analysis did not show any clear pattern of structure since there is a high variability in both the size and the number of introns present. However, when we compared the amino acid sequence of 23 PME genes from higher plants and a PME gene from E. chrysanthemi (Figure 3 a, b, c, d), the analysis highlighted a large region in common for most of the plant PME genes: GPXKHQAVALR. Also, we noted that it is located most of the time at the same place with respect to the presence of the first intron. Experiments of site directed mutagenesis with a PME gene from Aspergillus niger strain 5344 had shown that there is an histidine residue essential for PME activity within the amino acid sequence HQAVA (Duwe and Khanh, 1996). From Figure 3 a, b, c, d, we can see that most of the PME enzymes from higher plants has the sequence HQAVA as well. This seems to suggest that this histidine residue can be playing an important role in the catalytic activity of the enzyme. Multiple sequence alignment failed to locate the sequence of HQAVA of Erwinia chrysanthemi PMEA or PMEB at the same location as plant PME’s. However, pair comparison between PMEU1 and PMEA or PMEB from Erwinia chrysanthemi correctly aligned the sequence HQAVA at the same position. Studies of the three-dimensional structure of Erwinia chrysanthemi pectin methylesterase (PME-A) support the presence of two aspartate and one arginine residues in the active site of the enzime (Jenkins et al. 2001) and not an histidine. However, some of the PME isoenzymes show an aspartate residue instead of histidine in the same site (Figure 3 a, b, c, d). We believe that the study of the possible involvement of either an histidine or an aspartate residues in the catalytic activity of PME from higher plants deserves further attention. Computer analysis of the PMEU1 genomic region showed that this sequence follows several features commonly present in other genes from higher eukaryotic organisms, as mentioned above. The phylogenetic analysis (Figure 4) had shown that this PME gene is not related with other PME genes isolated from the tomato genome (Harriman et al. 1991). Rather, from Figure 4, we can see that PMEU1 is more related to two genes from Arabidopsis thaliana (AtPME2 and AtPME3) and two genes from Citrus sinensis (PECS-1.1 and PECS-1.2). Efforts to find a correlation between relatedness of the PME genes and pattern of expression were not succesful. However, the finding just mentioned further support that the cloned PME gene described in this work belong to a entirely novel type of PME gene from tomato. Experiments carried out in our lab with tobacco transgenic plants overexpressing the PMEU1 gene and tomato plant with lower levels of this gene did not produce a change in the plant phenotype that could be give us an insight as to what is the physiological role of the PMEU1 gene. Therefore, we decided to computer analyzed the PMEU1 promoter sequence to look for DNA boxes or elements with known function, in search for insights as to what can be the physiological role of this PMEU1 gene. In Figure 5 (a, b), it is presented the sequence of the DNA regulatory region of the PMEU1 gene. We are not sure of having the complete genomic sequence of the PMEU1 gene for two reasons: the DNA segment of the PMEU1 gene was located toward the 5´ end of the tomato genomic DNA carried by the isolated phage (Figure 1). Further, computer analysis of the PMEU1 promoter 5´ end region failed to find elements known to exist toward the 3´ end of the gene coding regions. However, considering the size of the largest sequence of a PME regulatory region published to date, 2.3 kb (Albani et al. 1991), it is quite possible that we almost had the entire PMEU1 regulatory region. Our efforts to isolate from the tomato genomic library the remaining segment of the PMEU1 regulatory region were largely unsuccessful. The computer analysis of the PMEU1 regulatory region showed the presence of both direct repeats and perfect inverted repeats. In Figure 5 (a, b), only the largest ones are shown. It is interesting that repeats 1 and 2, which are only separated by one base pairs appears to come from only one repeat in which a mutation took place, splitting this long repeats into two shorter ones. Also, some of the largest perfect inverted repeats are present inside of the largest direct repeats. It can be interested to test whether this repeats belong to the PMEU1 promoter or they are part of the intergenic region of the plant genome which is known to contain repeat sequences. However, the possible role if any of these repeats remains to be elucidated. We also located two sequences identical to cis-acting elements found in the phytochrome A3 promoter (Bruce et al. 1991). Also, it showed two more sequences similar to known cis-acting elements regulated by light. From here, the possible regulation of this PMEU1 gene by light deserves further attention. We also located a sequence similar to a known abscisic acid responsive element, close to the transcription start site (Figure 5 (a, b)). The phytohormone ABA had been related to the abscission phenomena en plants (Label et al. 1994; Aneju et al. 1999) and to the plant responses to abiotic stress in plant (Zhu, 2001). One of the genes encoding a pectin methylesterase isolated from Citrus sinenis was shown to be up-regulated in abscission zones of leaves (Nairn et al. 1998). Currently, experiments in our laboratory are being carried out to test the possible role of the gene PMEU1 in the plant responses to light, abscission and abiotic stress, however, a possible function for the PMEU1 gene in these phenomena is still matter of controversy. With the goal to demonstrate that the 5´ flanking region of the PMEU1 genomic clone correspond with an active regulatory region, and to find the smallest size of the region able to direct transcription, we created transgenic tobacco plants expressing different constructs in which the uidA gene, encoding the enzyme β-glucuronidase, is being regulated by different regions of the PMEU1 promoter. In Figure 6, it is shown the results of analyzing the β-glucuronidase activity of around 50 independent transformed tobacco plants. From the figure, it is clear that by reducing the size of the promoter, its transcriptional activity is also reduced. As can be seen, even 267 bp of the PMEU1 regulatory region is transcriptionally active. This means that we did not reach the lower limit where the promoter loose completely its transcriptional activity, although a large reduction was accomplished. In contrast, it was reported that a truncated piece of 440 bp of a flax PME promoter (Lupme3) lost completely the ability to drive transcription of a reporter gene (Roger et al. 2001). The results of GUS activity in leaf tissue are supported by the histochemical staining analysis in which the transgenic plants showed weaker activity in the parenchyma tissue surrounding the leaf vascular tissue with decrease in the promoter size (data not shown). The change in transcriptional activity among the different sizes of promoter is of 6 fold when comparing the 0.267 kb. with the 1.306 kb. and 4 fold when comparing the 1.306 kb. with the 2.59 kb. There is a difference of 1.03 kb between 0.267 kb and 1.306 kb and 1.29 kb between 1.306 kb and 2.59 kb. The differences in sizes are similar and still the variation in activity is higher between the 0.267 Kb. and 1.306 kb which means that perhaps there are stronger enhancer element(s) in the promoter region closest to the ATG. Overall, we obtained up to 95% in reduction of PMEU1 promoter transcriptional activity with the construct including 0.267 kb of PMEU1 promoter. Reduction of the promoter size which brings an associated reduction in promoter activity as measured with a reporter gene had been found in deletion studies of other promoters (Darasiela et al. 1996; Royo et al. 1996), however, sometimes smaller pieces are able to drive higher levels of reporter gene activity in general (Canevascini et al. 1996) or at some specific tissues (Royo et al. 1996). The standard deviation of the parameter is indicating a very high variability which is most likely due to the presence of multiple copies in the genome of the different transformants (not determined), dissimilarities in the physiological status among the leaf tissues used and to the position effect (Wilson et al. 1990). This result is alike with studies reported earlier, in which a high variability among independent transformants was also found in liquid cell cultures expressing the GUS gene under the manopine synthase (Peach and Velten, 1991). Also, tobacco cells stably transformed with a chimeric construct in which the CaMV35S was driving the expression of GUS, showed a standard deviation three times higher than average for the GUS specific activity parameter (Allen et al. 1993). The average of GUS activity in tobacco leaf for the construct harboring the 2.59 kb of promoter size was 324.334 pMoles of MU/min/mg protein (Figure 7). This activity is similar to the one reported earlier for tobacco (Nicotiana tabacum var Samsun) leaf of about the same size used in this work, harboring GUS (uidA gene) under the control of the cauliflower mosaic virus 35S promoter: 321 pMoles/min/mg protein (Jefferson et al. 1987). This result suggest that PMEU1 promoter is as strong as the CaMV35S which in turn indicates its usefulness in overexpressing proteins in plants. We also studied the expression of the three constructs in the three main plant tissues: root, stem and leaf of six independent transformants (Figure 7). It is clear from the graph that the three constructs showed the same pattern already observed for leaf tissue. However, we recorded 1.7 and 8 fold PMEU1 transcriptional activity for stem and root, respectively. This suggest a difference in the strength of the enhancer elements present in the PMEU1 promoter depending upon the type of plant tissue. These results also suggest that the enhancer element(s) are active en several differentiated tissues and are not specific for leaf tissue. These results are supported by the GUS histochemical staining in which the transgenic plants harboring the construct including the smallest promoter region showed weaker activity in the parenchyma tissue surrounding the vascular tissue as compared with tissues of transgenic plants expressing the construct with the highest promoter region (data not shown). These findings are in contrast with deletion studies of other promoter in which it was found that for specific tissues, smaller pieces of the regulatory regions are able to direct higher values of reporter gene enzymatic activity (Royo et al. 1996). Deletion studies of the PMEU1 promoter could be of significant insight to locate this putative enhancer elements. However, stronger experimental evidences are needed to probe their presence in the PMEU1 promoter region. In summary, we had isolated an entirely new gene encoding a pectin methylesterase isozyme from the tomato genome which is represented by a single copy. It shows an ubiquitous pattern of expression, in contrast with the tissue specific gene isolated earlier from tomato. Analysis of its promoter region suggest several potential function for this gene and we believe that further analysis of this gene will bring new insights to understand better the physiological role of the pectin methylesterase enzyme. References
Note: Electronic Journal of Biotechnology is not responsible if on-line references cited on manuscripts are not available any more after the date of publication. Supported by UNESCO / MIRCEN network © 2004 by Pontificia Universidad Católica de Valparaíso -- Chile The following images related to this document are available:Photo images[ej04003f4.jpg] [ej04003f7.jpg] [ej04003f1.jpg] [ej04003f5a.jpg] [ej04003f3a.jpg] [ej04003f2.jpg] [ej04003f3c.jpg] [ej04003f5b.jpg] [ej04003f3d.jpg] [ej04003f6.jpg] [ej04003f3b.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}