 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Electronic Journal of Biotechnology, Vol. 7 No. 1, April 15, 2004 Isolation of adh mutant of Lactobacillus rhamnosus for production of L(+) Lactic acid Niju Narayanan1, Pradip K. Roychoudhury2, Aradhana Srivastava*3 1Department of Biochemical Engineering and Biotechnology,
Indian Institute of Technology Delhi,
Hauz Khas, New Delhi – 110016, India
Tel: 91 11 26596192
Fax: 91 11 26582282
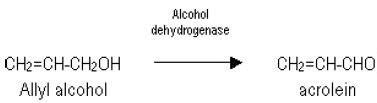
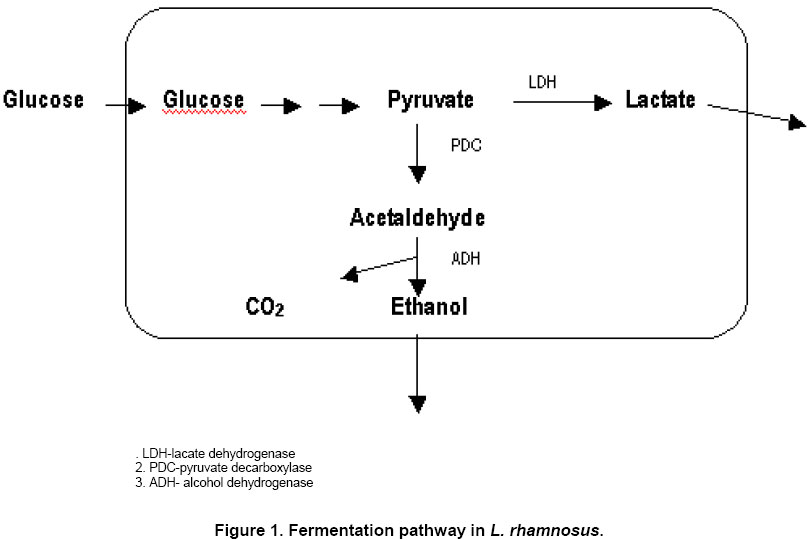
E-mail: nkniju@hotmail.com Financial support: research project supported by IIT Delhi. Received March 25, 2003 / Code Number: ej04009 Abstract Lactobacillus rhamnosus, a facultative anaerobe, which produces L (+) lactic acid and ethanol under anaerobic conditions, is used in the present study. An adh- mutant of Lactobacillus rhamnosus MTCC 1408, was developed by chemical mutagenesis, which could produce pure L(+) lactic acid as the only product. Batch fermentation kinetics of the wild type and the mutant strain were studied in glucose-yeast extract medium under conditions of temperature 40ºC and pH 6.2 anaerobically. The biomass yield was similar in both wild type and mutant strains, however lactic acid yield increased by 6.6%. A chemically defined media was optimized for supplementation of succinate, acetate and citrate for better biomass formation using single variable optimisation. It was further optimised for varying concentrations of vitamins, amino acids and trace metals by response surface method. The batch biomass yield (0.1g/g) and lactic acid yield (0.88g/g) in the optimised chemically defined media were similar to those obtained in the glucose-yeast extract medium. Keywords: anaerobic fermentation, ethanol, media optimization, mutagenesis, response surface method. L(+) lactic acid production is gaining interest due to its applications in food and pharmaceutical industries (Harrig, 1989; Datta, 1995). Most homofermentative organisms produce both the stereoisomeric forms of lactic acid. Heterofermentative organisms may produce lactic acid along with other metabolites like ethanol, acetate, formate etc., anaerobically. The purification of one form of lactic acid from its racemic mixture is very difficult, which may involve expensive chromatography techniques. The most suitable way is to select the L(+) producer organism or use genetically modified strains. A few attempts have been made to improve and modify the production of L(+) lactic acid by metabolic engineering in Lactobacillus species. The ldhD gene in L. helveticus was inactivated (Bhowmik et al. 1993). A two-fold increase in the amount of L(+) lactic acid was observed which restored the amount of total lactic acid to the level in the wild type strain. The ldhL gene was overexpressed in L. plantarum by increasing its copy number (Ferain et al. 1994). This increased the activity of L-lactate dehydrogenase (L-LDH) but was found ineffective for L(+) lactic acid production. The total lactic acid production was not affected by the inactivation of ldhD gene or inactivating the ldhL gene. In another study, two ldhD- strains of L. helveticus were constructed by deleting the promoter region of the ldhDgene in the first construct and by replacing the structural gene of ldhD gene by ldhL gene of the same species in the second construct (Nikkila et al. 2000). This has not only increased the enzyme activity of L-LDH by 53% and 93% respectively, but also enhanced L(+) lactic acid production. However, they could only produce the lactic acid rich in L(+) form, but there was a need to separate L(+) lactic acid from its D(-) form to facilitate its usage for medical field. The presence of D(-) lactic avid is interfering in the L(+) lactic acid polymer formation which is applied for medical purposes. In Lactococcus lactis, ldhLgene is located as a part of the lac operon. An increase in copy number of the plasmid containing the whole operon resulted in a slight increase in lactic acid production (Llanos et al. 1992). The ldhD gene of L. johnsonii was altered by an 8-bp deletion. The D-LDH enzyme activity was completely absent in the mutant strain and the remaining L-LDH enzyme activity rerouted the carbon flux to L-lactate with a marginal increase in the secondary end products (Lapierre et al. 1999). Many of the genetically modified homofermentative lactic acid bacteria were unable to produce pure L(+) form of lactic acid. The present studies were planned to produce L(+) lactic acid as the only product using the heterofermentative organism Lactobacillus rhamnosus. This organism produces ethanol along with L(+) lactic acid under anaerobic conditions (Figure 1). For this purpose adh- mutant of L. rhamnosus was developed by chemical mutagenesis. Its behaviour was examined in batch fermentation. The advantage of this method is the pure L(+) form of lactic acid production as the sole product. The downstream processing for the separation of pure L(+) lactic acid from the fermentation broth can be made easier if a defined media is used. The concentration of amino acids, vitamins and trace metals solution were optimised by experimental design and response surface method and their batch kinetics were observed in the optimised media. Materials and Methods Microorganism and culture condition The bacterial strain Lactobacillus rhamnosus MTCC 1408 (obtained from Institute of Microbial Technology, Chandigarh) is used. Stock cultures of the bacteria were stored in MRS medium as 15% glycerol stock at -80ºC (as instructed by Institute of Microbial Technology, Chandigarh). The medium used in the 2 % (w/v) agar plates, precultures, and batch fermentations contained 100 g/l glucose, 30 g/l yeast extract, 0.2 g/l KH2PO4, 0.2 g/l K2HPO4, 0.3 g/l MgSO4.7H2O, 0.03 g/l MnSO4.H2O, 0.03 g/l FeSO4, 2 g/l Sodium succinate, 2 g/l Sodium sulfate. The pH of the medium was adjusted at 6.2 by adding 2N NaOH/ 2N HCl before sterilization (Srivastava et al. 1992). L. rhamnosus was anaerobically grown on 2% (w/v) agar plates at 40ºC for 24 hrs, and then kept at 4ºC. Initial preculture for bioreactor cultivations was grown in glucose-yeast extract medium at temperature 40ºC for 24 hrs inside anaerobic jars. 2% (v/v) inoculum was used from this culture to prepare the successive preculture. This was further used for inoculating the fermentor. Bioreactor cultivations were performed in a 3.0 l glass fermentor (KLF 2000, Bioengineering AG, Wald Switzerland) using 2 l working volume. The fermentor was equipped with all monitors and controllers for pH, temperature and dissolved oxygen. An agitation speed of 300 rpm was maintained with the help of agitator fitted with two flat blade impellers. Anaerobic conditions were maintained by sparging nitrogen gas before inoculation and intermittently during the course of fermentation. The pH was maintained at 6.2 by automatic addition of 4N NaOH/ 4N H2SO4 during lactic acid fermentation. The schematic diagram of the set up is given in Figure 2. Actively growing cells from the second preculture in glucose-yeast extract medium, were collected and centrifuged to separate cells from supernatant. The cells were then washed with 0.1 M citrate buffer (pH 5.5), resuspended in 0.1 M citrate buffer (pH 5.5), and then mutagenised by exposing these cells to N- methyl, N'-nitro, N-nitrosoguanidine (MNNG, 50 mg/ml) for 30 minutes at 28ºC. Thereafter, cells were washed with 0.1 M phosphate buffer (pH 7.0). N- methyl, N'-nitro, N-nitrosoguanidine, breaks down at pH 7.0. The washed cells were suspended in glucose-yeast extract medium and spread onto agar plates 0.6% (v/v) allyl alcohol. This compound inhibits wild type cells since cells with functional alcohol dehydrogenase activity convert allyl alcohol to the toxic compound acrolein (Pulci et al. 1993).
Single colonies obtained on plates were repeatedly transferred to agar plates containing 0.6% (v/v) allyl alcohol to get a stable mutant. Analytical methodsOptical density of the culture was measured at 600 nm with a spectrophotometer (spekol 1100, analytik jena, Germany) The dry cell weight was determined by centrifuging the culture broth to separate cells from supernatant, then washing with 0.85% NaCl solution and drying in preweighed aluminium foil cups at 70ºC till constant weight. One optical density was found to be equivalent to 0.34 g dry cell weight per litre. Concentrations of glucose in samples were determined by Dinitro Salycilic Acid method (Miller, 1959). Total lactic acid was determined colorimetrically using p-phenyl phenol (Barker and Summerson, 1941). L(+) lactic acid concentration were analysed spectrophotometrically at 340 nm using L-lactate dehydrogenase enzyme (L-LDH). The assay mixture containing 0.6 M glycine, 0.47 M hydrazine, 35 mg/ml NAD, 13.6mg/ml L-LDH and the appropriately diluted test samples were incubated at 30ºC for 90 min before taking optical density at 340 nm. The concentration of ethanol, acetate, formate were determined by a gas chromatograph (AIMIL, Nucon, India, Series 5765) equipped with Chromosorb 101 column using nitrogen as the carrier gas and a mixture of hydrogen and oxygen gas to sustain the flame. The detector, injector and oven temperature were maintained at 200ºC, 195ºC and 180ºC respectively. A gas flow rate of 35 ml/min was maintained. A glucose limited defined media (composition obtained from Centre for Process Biotechnology, DTU, Denmark) was taken as the basis. Saline Solution I (containing NH4Cl 0.142 g/l and K2SO4 0.059 g/l), Saline Solution II (containing KH2PO4 0.197 g/l, NaCl 3.247 g/l, MgCl2 0.118 g/l and FeSO4 3.09x10-3 g/l), trace metal solution (containing (NH4)6(MO7)24 4.11x10-6 g/l, H3BO3 27.44 x10-6 g/l,CoCl2 7.9 x10-6 g/l, CuSO4 2.88 x10-6 g/l,MnCl2 13.9 x10-6 g/l, ZnSO4 3.1 x10-6 g/l andCaCl2 8.17 x10-6 g/l) and glucose (10 g/l) were sterilised separately at 121ºC for 20 min. Amino acid solution (containing L-alanine 0.168 g/l, L-arginine 2.71 g/l, L-asparagine .133 g/l, L-cysteine 0.647 g/l, L-glutamate 0.163 g/l, L-glutamine 0.341 g/l, L-glycine 0.226 g/l, L-histidine 0.052 g/l, L-isoleucine 0.35 g/l, L-leucine 0.350 g/l, L-lysine 0.284 g/l, L-methionine 0.116 g/l, L-phenylalanine 0.220 g/l, L-proline 0.128 g/l, L-serine 1.354 g/l, L-threonine 0.224 g/l, L-tryptophan 0.227 g/l, L-tyrosine 0.06 g/l and L-valine 0.234 g/l) and vitamin solution (containing biotin 0.109x10-3 g/l, pyridoxal 2.28 x10-3 g/l, folic acid 1.128 x10-3 g/l, niacinamide1.083 x10-3 g/l , thiamine HCl 1.118 x10-3 g/l, calcium pantothenate 0.528 x10-3 g/l) were added by sterile filtration using 0.2 m filter. In this medium, 0.2 g/l of MnSO4 was added as according to the composition of MRS medium composition recommended by Institute of Microbial Technology, Chandigarh. The experiments were carried out in 500 ml flasks with 200 ml medium, inoculated with 2 ml culture from preculture II, and incubated at 40ºC for 48 hrs under anaerobic conditions without shaking. The various combinations of acetate, citrate and succinate concentrations investigated are shown in Table 3.
Statistical experimental design A Box Behnkehn experimental design with three variables, amino acids, vitamins, trace metals solution and the two parameters or responses, lactic acid and biomass were used which resulted in a total of 20 experiments. The real and coded values of the variables are shown in Table 2. Design Expert version 5.5 (Statease Inc, MN, USA) software was used for statistical experimental design, analysis of the results and drawing of the contour plots. The model was fitted by multiple linear regression and selected by examining the ‘Lack of Fit tests’ in which low ‘ROOT MSE’ and ‘PRESS’ values describe the fitness of the model.
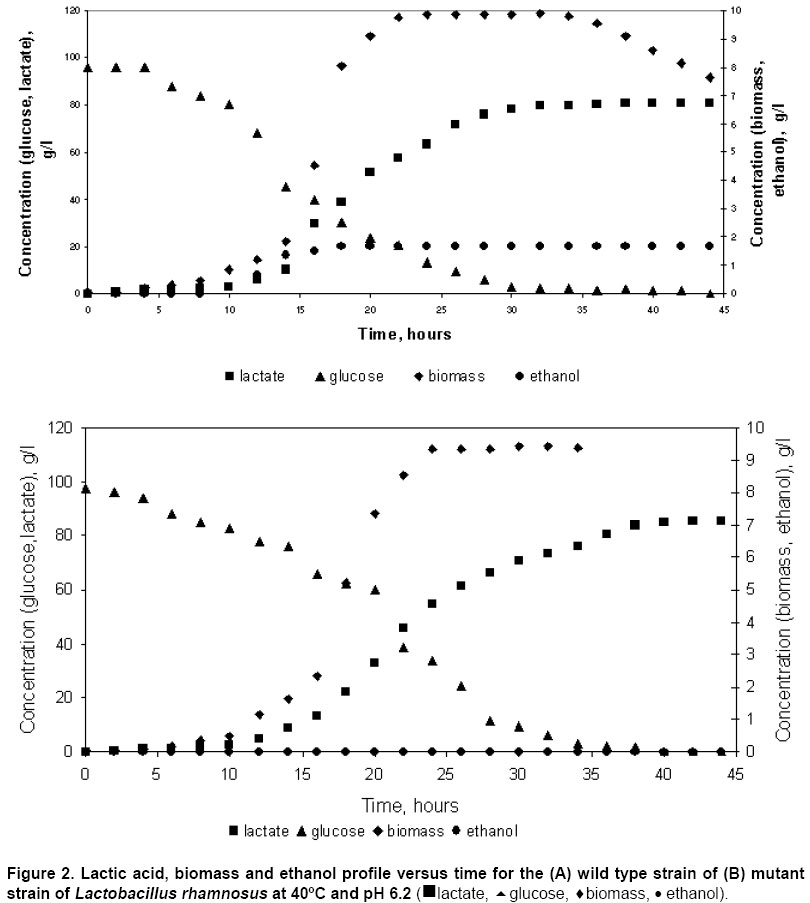
Physiology of L. rhamnosus MTCC 1408 wild type The batch fermentation kinetics of the wild type strain of L. rhamnosus was studied at 40ºC and pH 6.2 under anaerobic conditions initial glucose concentration of 100 g/l. Cells grew exponentially up to 18 hrs and the cell growth ceased after 24 hrs. The fermentation profiles of biomass, glucose, lactic acid and ethanol are shown in Figure 2 (A). Lactic acid was produced to 50 g/l during the exponential growth of the cells but it continued up to 34 hrs to a final concentration of 80.3 g/l. After 24 hrs, the lactic acid production rate is almost constant. Lactic acid production is known to be growth associated and non-growth associated production. The production of ethanol has also shown the similar pattern. There observed a rapid increase until 24 hrs, and thereafter constant rate of production is obtained. Ethanol production also ceased with cell growth at a final concentration of 4.1 g/l. The biomass concentration was found to be 9.86 g/l. The specific growth rate was observed to be 0.31 h-1. The anaerobic glucose metabolism in L. rhamnosus MTCC 1408 produces lactic acid and ethanol as the fermentation product. Other products like pyruvate, acetate, acetaldehyde were found absent in the fermentation samples which is clearly observed in gas chromatograph. This has suggested us that selection of L. rhamnosus to be used for anaerobic L(+) lactic acid production is very suitable. The anaerobic batch fermentation profiles have also shown the little substrate diversion for ethanol (another metabolite of anaerobic fermentation). Although, major portion of this produced ethanol can be separated by physical purification/separation techniques, but for diverting the whole carbon substrate to L(+) lactic acid via anaerobic fermentation, the development of adh- mutant was required to produce. Moreover, the presence of even slightest amount of ethanol is hampering the polymerisation of L(+) lactic acid which was the set for the present work. Cells were exposed to N-methyl, N'-nitro, N-nitrosoguanidine (MNNG) solution of 100 mg/ml, 75 mg/ml and 50 mg/ml to develop adh- mutants that cannot produce ethanol. When 100 mg/ml MNNG was used for 20, 30 and 40 min, 3 colonies were obtained on allyl alcohol plates. These colonies were stable upto two transfers on allyl alcohol plates. When 75 mg/ml MNNG was used for 30 min, 3 colonies were obtained on allyl alcohol containing plates. These colonies also lost their ability to grow on allyl alcohol containing agar plates after 2 or 3 transfers on agar plates containing allyl alcohol. With 50 mg/ml MNNG, cells from broth obtained at different fermentation time (exhibiting optical density 0.63, 0.81, and 1.24), were exposed for different time intervals (20, 30 and 40 min). From this study, 36 mutants were isolated, out of which 12 mutants were able to grow on allyl alcohol plates after 6 transfers. When the growth of the 12 stable mutants were investigated for their growth in flasks, the biomass obtained after 48 hrs showed a difference. Lactobacillus rhamnosus NN5 that showed the highest biomass concentration obtained after 48 hrs was selected and used for further studies. The results are summarised in Table 1.
Physiology of L. rhamnosus NN5 mutant strain The batch kinetics of the mutant strain L. rhamnosus NN5 was studied in a bioreactor operating at similar conditions that used in batch culture with wild type strain. Minor differences in the growth profiles of the mutant strain and that of the wild type were found. The specific growth rate had decreased from 0.31 h-1 for the wild type strain to 0.28 h-1 for the mutant strain. The wild type strain produced 4.1 g/l ethanol. No yield of ethanol was observed in fermentation by the mutant strain. The lactic acid concentration increased from 80.3 g/l for the wild type strain to 85.6 g/l for the mutant strain. However the overall productivity for lactic acid dropped from 2.38 g/l/h for the wild type strain to 2.14 g/l/h for the mutant strain. This observation could be due to the decrease in the observed maximum specific growth rate. Inspite of a decrease observed in lactic acid productivity, the lactic acid yield was enhanced by 6.6% clearly indicating the substrate amount which was diverting for the ethanol in wild type, being also utilized for L(+) lactate formation in the mutant strain. The fermentation profiles of biomass, glucose, lactic acid and ethanol are shown in Figure 2 (B). The comparison of yields and productivities for the wild type and the mutant in the glucose yeast extract medium is given in Table 5.
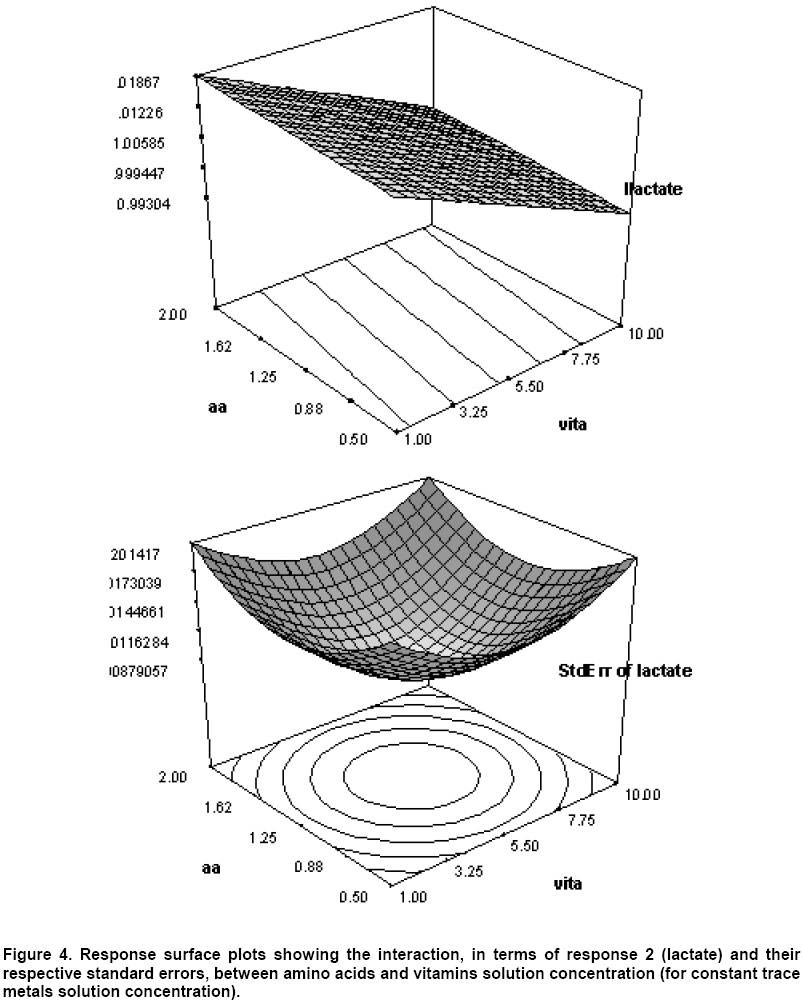
Although, the mutant showed the capability to produce L(+) lactate as the only product in complex glucose limited medium, but the proteins present due to unutilized yeast extract were found inhibitory for L(+) lactic acid polymerization. Therefore, further efforts were required to be made in order to minimize the protein contents in fermented medium. Optimization of glucose limited defined medium by response surface method In order to find the optimal concentrations of amino acids, vitamins and trace metals in the glucose limited defined media for lactic acid production, a response surface method was applied for the mutant strain L. rhamnosus NN5. These results of the experimental design are summarized in Table 3. These results were fitted to the model by linear regression. A response surface for cell dry weight of L. rhamnosus as a function of amino acids, vitamins and trace metals is presented in Figure 4. The ‘ROOT MSE’ and ‘PRESS’ values were 0.32 and 1.95 respectively. Figure 4 shows the response surface for the model predicting lactic acid formation by L. rhamnosus as function of amino acids, trace metals and vitamins. In this model also, ‘ROOT MSE’ and ‘PRESS’ (0.036 and 0.03 respectively) are lower than the quadratic model. Both lactic acid and biomass have been found to be a linear function of amino acids, vitamins and trace metals. The intermediate metabolites of tricarboxylic acid cycle are responsible for the formation of amino acids, vitamins, purines and pyrimidines. During anaerobic fermentation pyruvate gets converted to lactic acid or ethanol and does not enter the TCA cycle. Therefore intermediates have to be added externally to account for the same. In order to investigate the effect of supplementation of succinate, acetate and citrate, flask cultures were conducted by adding one component and excluding the other two or by adding two and excluding one of them. The optical density of the culture after 48 hrs and 60 hrs are shown in Table 4. The addition of citrate showed the cell concentration was the highest followed by acetate and succinate showed the lowest cell concentration. Also acetate with citrate (each at 1 g/l concentration) gave the same highest optical density of culture as that of acetate, citrate, and succinate (each at 1 g/l concentration). Therefore the concentration of acetate with citrate (each at 1 g/l concentration) was taken in the medium for subsequent experiments.
Fermentation physiology of mutant strain in optimised glucose limited defined medium To investigate the performance of the mutant strain of L. rhamnosus in the optimized medium, fermentation operation was carried out using initial glucose concentration of 100 g/l. The fermentation run was performed at controlled pH of 6.2. The fermentation profiles for biomass, glucose and lactic acid concentration (g/l) are shown in Figure 6. Cells reached their exponential growth phase after 4 hrs and reached their stationary phase by 20 hrs. Lactic acid was produced to a concentration of around 45 g/l with cell growth but continued to be produced to a final concentration of 85.6 g/l up to 45 hrs. The batch kinetics of the mutant strain in the optimized synthetic medium and the glucose yeast extract medium are comparable. For the glucose yeast extract medium, the rate of substrate consumption has been observed to be maximum during the exponential phase of growth, but wherein lactic acid was also being produced exponentially for the mutant and the wild type strain. Ethanol was also being produced by the wild type strain during the exponential growth of the cells, whereas in the optimized defined medium, a steady consumption of glucose was observed. Although, the specific cell growth rate in this medium (0.23 h-1) was lower than the case with the complex medium (0.28 h-1), the maximum lactic acid concentration of 85.6 g/l could be obtained. The comparison of yields and productivities for the mutant strain in the optimized synthetic medium are given in Table 5. The biomass and the lactic acid yields were similar in the two media for the mutant strain. However, the volumetric productivity is found to be 8.9% less in the case of optimised medium than the glucose-yeast extract medium. This was observed due to the lowering of observed maximum specific growth rate in defined glucose limited medium. The mutant L. rhamnosus NN5 obtained by chemical mutagenesis (MNNG) is able to perform stably in batch fermentation without reverting back to the wild type. The observed maximum specific growth rate of the mutant strain was 0.28 h-1 as compared to 0.31 h-1 obtained for wild type strain in glucose-yeast extract medium. A small decrease in lactate productivity from 2.38 g/l/h for the wild type strain to 2.14 g/l/h for the mutant strain was observed in glucose-yeast extract medium. An optimal defined medium was obtained using response surface method. The adh- mutant was able to produce L(+) lactic acid as the only product with a yield of 0.88 g/g in the optimised glucose limited defined medium. L(+) lactic acid is a monomer for the biodegradable poly L-lactate polymer industry. The production of L(+) lactic acid can further be improved by various process strategies for fermentation and downstream processing (Srivastava et al. 1999).
Note: Electronic Journal of Biotechnology is not responsible if on-line references cited on manuscripts are not available any more after the date of publication. Supported by UNESCO / MIRCEN network © 2004 by Pontificia Universidad Católica de Valparaíso -- Chile The following images related to this document are available:Photo images[ej04009f2a-b.jpg] [ej04009f4.jpg] [ej04009f1.jpg] [ej04009f5.jpg] [ej04009f3.jpg] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||

{kind=link}
{kind=link}
{kind=link}