 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Electronic Journal of Biotechnology, Vol. 7, No. 2, August, 2004 RESEARCH ARTICLE Catalytical properties of
N-glycosylated Gluconacetobacter diazotrophicus levansucrase produced
in yeast
Luis Enrique Trujillo Toledo*1, Raúl Gómez Riera2, Alexander Banguela Castillo3, Melvis Soto Romero4, Juan Gabriel Arrieta Sosa5, Lázaro Hernández García6, 1Division Plantas,
Laboratorio Interacciones Planta-Microorganismos,
Centro de Ingeniería Genética y Biotecnología,
P.O. BOX 6162,
C. Habana, Cuba
Tel: 53 7 2716967/6413 ext. 3117
Fax: 53 7 331779
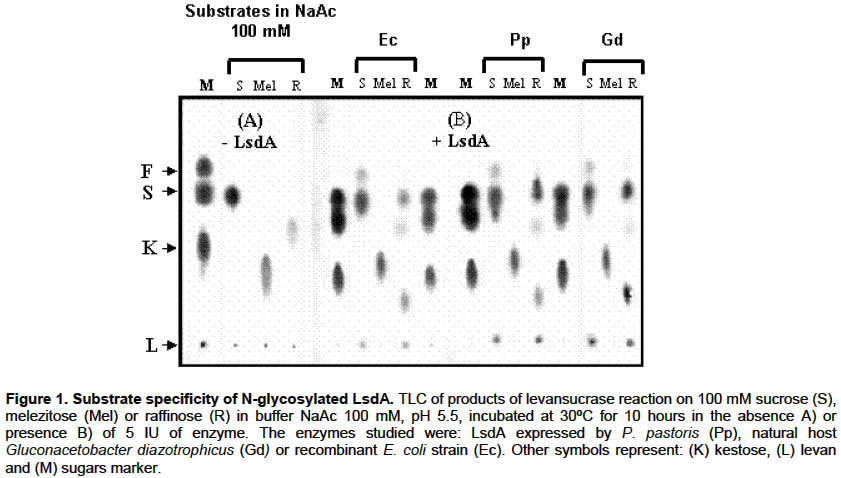
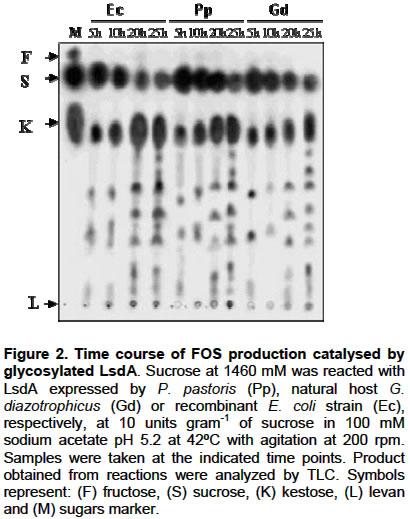
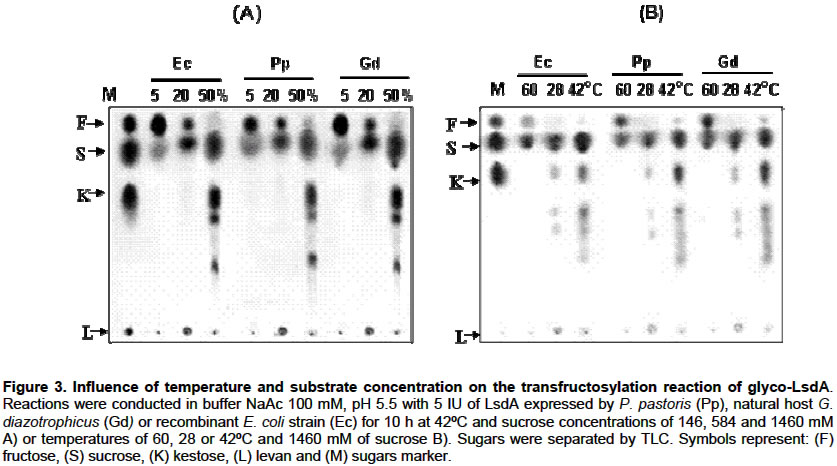
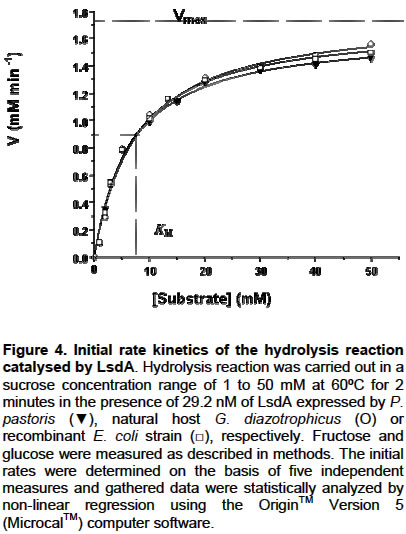
E-mail: luis.trujillo@cigb.edu.cu Financial support: This research was supported by a project grant of the Cuban government # 3031254. Received January 7, 2004 / Code Number: ej03013 ABSTRACT The influence of N-glycosylation on the kinetic and catalytical properties of a bacterial fructosyltransferase (LsdA) produced in Pichia pastoris was studied. The glycosylated enzyme behaved similarly to non-glycosylated LsdA when substrate specificity, fructo-oligosaccharide (FOS) production, sucrose hydrolysis or levan formation reactions were carried out under different experimental conditions. The kinetic parameters for native or yeast-expressed LsdA determined at 60ºC, condition for the highest hydrolytic activity, followed a conventional Michaelis-Menten kinetics. Synthase activity of this levansucrase increased in water-restricted environments by addition of salt or organic solvent to the reaction mixtures. Keywords: catalytical properties, fructo-oligosaccharides, fructosyltransferase, levansucrase, N-glycosylation, Pichia pastoris. Abbreviations: ARTICLE Gluconacetobacter diazotrophicus levansucrase (LsdA) catalyses four types of reactions: 1) hydrolysis, where sucrose splits into fructose and glucose, 2) Fructo-oligosaccharides (FOS) production by transferring the fructosyl residue from a donor to an acceptor sucrose molecule forming b(2->1) links, 3) polymerisation, where fructosyl residues are b(2->6) linked to a fructan polymer and 4) exchange reaction where the fructosyl residue is transferred to a glucose molecule to form sucrose, (Hernández et al. 1995). FOS consumption benefits human and animal health by increasing bifidobacteria population in the intestinal track (Simmering and Blaut, 2001). Among them, the most attractive, 1-kestose, is produced with high yield by LsdA action on sucrose (Hernández et al. 1995; Támbara et al. 1999). Mature native LsdA has a N-terminal pyroglutamic acid and a disulfide bridge as unique modifications (Betancourt et al. 1999) however; expression of the lsdA gene in Pichia pastoris yielded an active but glycosylated enzyme (Trujillo et al. 2001; Trujillo et al. 2002). Structure and functional significance of glycosylation for different proteins has been increasingly appreciated (Patel et al. 1997; Montesino et al. 1998; Gemmill et al. 1999). Since LsdA constitutes a promising candidate for industrial production of FOS, we investigated the catalytic and kinetic properties of the glycosylated enzyme: i- substrate specificity, ii- product profile as a result of transfructosylations reactions on sucrose conducted in both, aqueous or water-restricted environments and iii- kinetic parameters for the hydrolysis reaction; in comparison with non-glycosylated LsdA expressed by its natural host G. diazotrophicus and by a recombinant E. coli strain. To our knowledge, there are no reports dealing with the catalytic and kinetic properties of a recombinant glycosylated levansucrase, in aqueous or water-restricted environments. MATERIAL AND METHODS Enzymes Native levansucrase used as standard in all the biochemical and kinetics experiments was purified from culture supernatants of G. diazotrophicus strain SRT4 (Hernández et al. 1995). The N-glycosylated LsdA was obtained from a recombinant P. pastoris as previously described (Trujillo et al. 2001; Trujillo et al. 2002). A recombinant E. coli strain harboring the plasmid pALS200, where the lsdA gene was fused at its 3’ end to the myc epitope and polyhistidine tag coding sequence placed under the transcriptional control of the nptII promoter, was also used as a recombinant non-glycosylated source of LsdA. Protein purification, quantification and enzyme assays Purification of recombinant LsdA myc-His6 produced in both, P. pastoris and E. coli was performed from culture supernatants and cell lysates respectively, by Immobilization-Metal Affinity Chromatography, using Ni-NTA agarose beads from QUIAGEN (Hilden, Germany). Total protein in culture supernatants, crude extracts and final purified preparations was determined as described by Bradford, 1976. Specific activity values of each enzymic preparation were calculated as the average of 5 different experiments. After ANOVA test, no significant differences were found between the analysed means. LsdA purity was determined by densitometric analysis of 12.5% polyacrylamide gels stained with Coomassie brilliant blue R-250 (Sigma, USA). Levansucrase activity was measured as the glucose released from sucrose using a glucose oxidase-peroxidase-coupled colorimetric kit (Sigma). One unit of LsdA is defined as the amount of enzyme releasing 1 µmol of glucose min-1 based on initial velocity measurements under the following conditions: 100 mM sucrose in 100 mM sodium acetate buffer pH 5.5, at 30ºC. The dinitrosalicylic acid (DNSA) test was also used to detect released glucose-fructose during hydrolisis reactions. Fructan detection and quantification Sugar samples (1 µl) were spotted on a Thin Layer Chromathography (TLC) foil (Merck KGaA, Darmstadt, Germany). TLC foils were run thrice in acetone:water (90:10) mixture and then, fructose–containing sugars were specifically stained with a urea-spray. This spray was prepared as follows: Urea (3 g) was dissolved in 100 ml of 1 M phosphoric acid in water-saturated butanol. Ethanol (5 ml) was added to eliminate the water phase which forms when the urea dissolves. TLC foils are then incubated at 100ºC for several minutes. Total sugars in the reaction mixtures were also analysed by High Pressure Liquid Chromatography (HPLC) using an Aminex HPX-87N column (matrix DVB styrene sodium, BIO-RAD) with a refractive index detector. A solution of 10 mM Na2SO4 was used as mobile phase at a flow rate of 0.5 ml min-1. Known amounts of sucrose, glucose, fructose and kestose were used as standars. RESULTS Throughout this paper we studied the catalytical properties of glycosylated LsdA produced in P. pastoris however, for subsequent comparison we also included as controls in our experiments both, the native enzyme from Gluconacetobacter diazotrophicus (Hernández et al. 1995) and a recombinant LsdA produced in Escherichia coli. These two control enzymes share in common lacking N-linked carbohydrates. The three LsdA preparations displayed 95-96% purity with specific activity values of 160 ±12 U mg-1 calculated as described in methods. Substrate specificity and FOS production by glycosylated LsdA Glycosylated LsdA produced in P. pastoris reacted on sucrose and raffinose [O-a-D-galactopyranosyl-(1->6)-O-a-D-glucopyranosyl-(1->2)-b-D-fructofuranosidose] to form free fructose and levan in a proportion similar to those of the native or E. coli–produced enzymes used as controls (Figure 1). However, the enzyme did not react on melezitose [O-a-D-glucopiranosyl-(1->3)-b-D-fructofuranosyl-(2->1)-a-D-glucopiranoside], in the same fashion than native and E. coli expressed LsdA (Figure 1). The time course of FOS production from sucrose indicated that the glycosylated enzyme converts efficiently sucrose to FOS, mainly 1-kestose (Figure 2). Dependence of transfructosylation reaction of glyco-LsdA on reaction conditions Qualitative analysis of the product profile obtained after incubation of the three enzymes under different reaction conditions showed that variations in temperature and sucrose concentration altered the product profiles of all the assayed enzymes in a similar way. (Figure 3 A, B). The highest levels of 1-kestose were produced at sucrose 50% (w/v, 1460 mM), whereas at the lowest substrate concentration (5%, 146 mM), the enzymes mainly hydrolyzed sucrose and consequently the 1-kestose levels were low (Figure 3A). The effect of temperature on the transfructosylation reaction catalyzed by the three enzymes was examined (Figure 3B). Fructose release was maximal at 60ºC indicating that the hydrolyzing activity of the assayed enzymes increased with temperature rise, while the highest conversion of sucrose to 1-kestose was reached at 42ºC. Levan formation was favoured at 28ºC. The three enzymes showed unaltered product profiles in the pH range from 5 to 8 (results not shown). The results presented in this section are in agreement to those previously reported for native G. diazotrophicus levansucrase (Támbara et al. 1999). Kinetics of hydrolysis reaction catalysed by glycosylated LsdA The influence of sucrose concentration on initial hydrolysis velocity rates (Vo) was evaluated. The chosen assay temperature of 60ºC and pH 5.5 corresponds to conditions of the highest hydrolytic activity for native and recombinant LsdA. Under these reaction conditions only released glucose and fructose were detected as reaction products after quantification of total sugars in the reaction mixtures. Statistical analysis of data gathered from this kinetic assay showed a correctly fit with a rectangular hyperbole as depicted in Figure 4, therefore obeying the conventional Michaelis-Menten kinetics for monosubstrate reactions where: VoG=VoF= Vmax [So] / Km + [So]. The VoG and VoF values correspond to initial velocity of glucose and fructose released during the hydrolysis reaction, meanwhile [So] is the initial sucrose concentration. The Michaelis constant (Km), maximal velocity (Vmax) and catalytical constant (kcat) values determined under these experimental conditions are listed in Table 1. We found no significant differences between the values of the above kinetic parameters for the three assayed enzymes as a result of its action on sucrose, indicating that N-oligosaccharides moieties of the P. pastoris produced enzyme affect neither catalytical velocity nor enzyme-substrate affinity. Results of kinetic studies conducted also at 30ºC did not fit a Michaelis-Menten behaviour (results not shown).
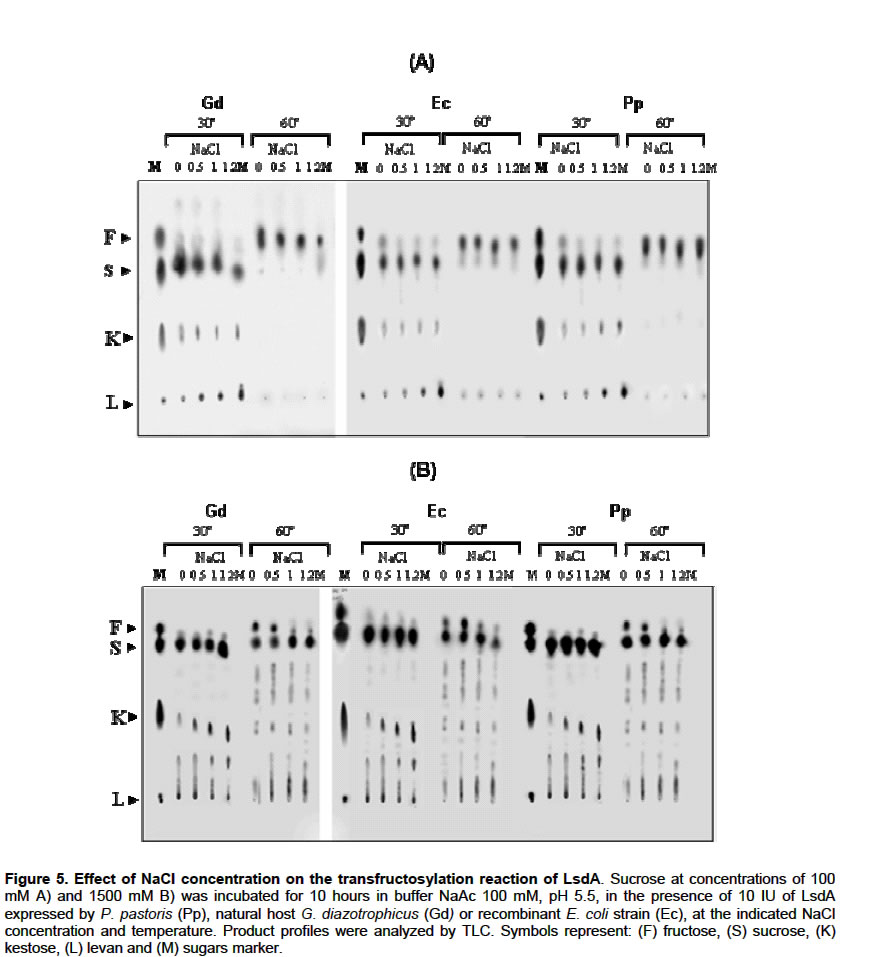
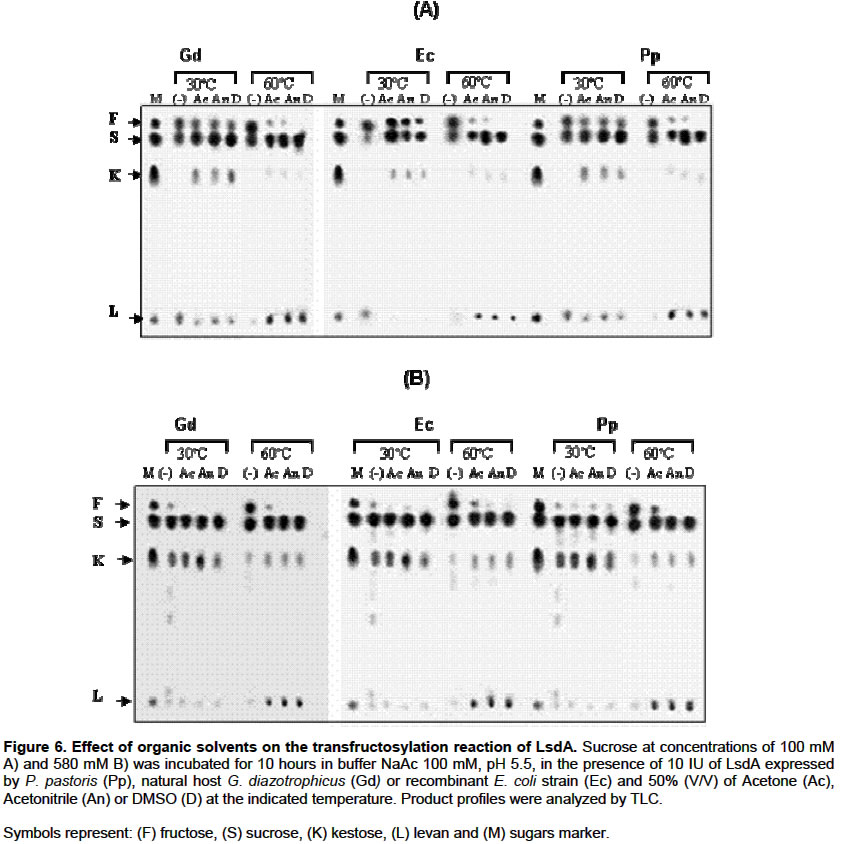
Catalytical properties of glycosylated LsdA in water–restricted environments Water participates in the reactions catalyzed by LsdA. There are no reports dealing with catalytic properties of LsdA, neither native nor recombinant in water-restricted environments. The equilibrium between hydrolase (glucose+fructose) and synthase (levan formation) activities was determined at 60ºC and 30ºC respectively, by progressively substituting water by addition of NaCl or different organic solvents into the enzymes reaction mixture, at different substrate concentrations. Figure 5A shows that levan formation at 30ºC increased with the increment of NaCl concentration from 0 to 1.2 M using sucrose 0.1 M as substrate. When substrate concentration was raised up to 1.5 M (Figure 5B), synthesis of 1-kestose but not polymerization was stimulated by NaCl concentration increments. Vigants and co-workers (1998) found that NaCl activated the polymerization reaction catalyzed by Zymomonas mobilis levansucrase in a way inversely proportional to the substrate concentration. These authors did not study FOS production under these conditions. At 60ºC, the amount of fructose liberated by LsdA decreased with increasing NaCl concentrations. Acetone, acetonitrile and dimethyl sulfoxide share the same property of being water miscible. When sucrose concentration was 0.1 M, the presence of these organic solvents at 30ºC stimulated 1-kestose syntheses with a very low rate of levan formation, while at 60ºC, levan synthesis was enhanced. Free fructose, as a result of the hydrolase activity of the enzymes, was only detected in the absence of solvents (Figure 6A, B). When the same reactions were conducted on sucrose 0.58 M at 30ºC, no remarkable differences were observed between the products profiles obtained in aqueous or anhydrous environments, however, at 60ºC the hydrolase activity was detected only in the reaction carried out in aqueous condition, while the presence of the organic solvents stimulated the synthase activity in all the assayed enzymes. DISCUSSION Foreign proteins are fortuitously glycosylated when secreted in yeast. This post-traslational modification is absent in native LsdA and other levansucrases due to their bacterial origin and could affect unpredictably the enzyme activity and stability. We studied the effect of N-glycosylation on the Gluconacetobacter diazotrophicus levansucrase (LsdA) recombinantly produced in Pichia pastoris. The predicted sequence of this recombinant enzyme contains four potential N-linked glycosylation sites, conforming to the general rule N-X-T/S where X is not proline however, the exact nature and location of the detected carbohydrate moiety have not yet been determined. We found no effects of glycosylation on substrate specificity, FOS production, sucrose hydrolysis or levan formation by LsdA ractions carried out under different conditions. By contrast, changes in substrate specificity and protein conformation have been reported in other recombinant fructosyltransferases produced in P. pastoris (Hochstrasser et al. 1998; Tibbot et al. 1998). The kinetic parameters for native or yeast-expressed LsdA determined at 60ºC, condition for the highest hydrolytic activity, followed a conventional Michaelis-Menten kinetics. The catalytic turnover rate displayed by the three assayed enzymes was approximately 10 times higher than that found for native LsdA at 30ºC (Hernández et al. 1995). The enzymes also showed higher affinity for sucrose at 60ºC (Km= 7.1 mM) than that displayed at 30ºC (Km=11.8 mM) previously described (Hernández et al. 1995). Interestingly, van Hijum and co-workers (2003) found that the Lactobacillus reuteri inulosucrase has lower affinity on sucrose during the hydrolysis reaction at 50ºC than at 20ºC. Glycosylated LsdA does not follow a monosubstrate type of kinetics at 30ºC. These findings result quite logical since at 60ºC the enzyme acts as a hydrolase, while at 28-30ºC the enzyme catalyzes both, hydrolysis and polymerization reactions (Figure 2B) hence, at this particular temperatures the enzyme behaviour adjust better to a bisubstrate than to a monosubstrate type of kinetics. All these results pointed out that the presence of N-linked oligosaccharides did not alter the catalytic properties of LsdA. According to 3D structural studies of levansucrases none of the four potential N-glycosylation sites are located in the active site of the ß-propeller architecture of LsdA precluding direct implications on catalysis (Meng and Futterer, 2003; Pons et al. 2004). Although the kinetics and catalytical properties studied herein were not affected by N-glycosylation, we reported in a previous work an increment in the thermal stability of glycosylated LsdA produced in P. pastoris when compared with the native enzyme (Trujillo et al. 2001). The stabilization role of carbohydrate moieties on glycoproteins has been described mainly for proteins naturally produced by fungi or yeasts (Wang and Hirs 1977; Patel et al. 1997; Boer et al. 2000). From the biotechnological standpoint is interesting the potential of most enzymes to vigorously work in organic solvents containing little or no water (Klibanov, 2003). We here show for the first time the possibility of modulating LsdA catalysis, including FOS production, by handling the concentration of organic solvents and temperature in the reaction media. Bacillus subtilis levansucrase is also active in water-restricted environments (Chambert and Petit-Glatron, 1989). Industrial FOS production, to date, depends chiefly on fungal enzymes (Yun, 1996). A deeper insight into the catalytical and kinetic properties of the P. pastoris-produced LsdA confirms that this recombinant glycosylated enzyme represents a promising candidate for mass production of FOS from sucrose in both aqueous or water-restricted environments. REFERENCES
Note: Electronic Journal of Biotechnology is not responsible if on-line references cited on manuscripts are not available any more after the date of publication. Supported by UNESCO / MIRCEN network Copyright 2004 by Universidad Católica de Valparaíso -- Chile The following images related to this document are available:Photo images[ej04013f5.jpg] [ej04013f3.jpg] [ej04013f4.jpg] [ej04013f6.jpg] [ej04013f2.jpg] [ej04013f1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}