 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
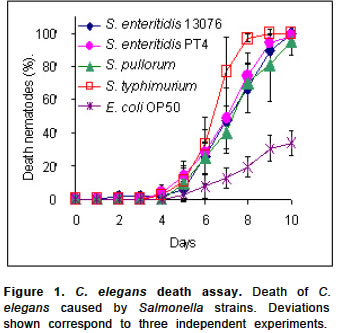
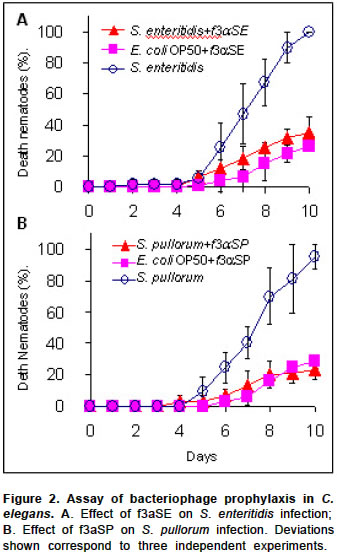
Electronic Journal of Biotechnology, Vol. 7, No. 2, August, 2004 SHORT COMMUNICATION Bacteriophage prophylaxis against Salmonella enteritidis and Salmonella pullorum using Caenorhabditis elegans as an assay system Javier Santander1, James Robeson*2 1Pontificia Universidad
Católica de Valparaíso,
Instituto de Biología, Laboratorio de Bacteriología,
Av. Brasil 2950 Valparaíso, Chile
E-mail: jrobeson@ucv.cl Financial support: Vice-rectoria de Investigación y Estudios Avanzados Pontificia Universidad Católica de Valparaíso. Project D.I.UCV 122.762.2001. Received November 13, 2003 / Code Number: ej04020 Keywords: animal model, bacteriophage prophylaxis, Caenorhabditis elegans, Salmonella. The nematode Caenorhabditis elegans has been used as animal model system to study bacterial pathogenesis. In this study we used C. elegans in a bacteriophage prophylaxis assay to test phage protection against infection by Salmonella enteritidis and S. pullorum. We found that pre-treatment of nematodes with bacteriophage that lyse S. enteritidis and S. pullorum results in enhanced survival of C. elegans, when challenged with these bacterial pathogens. It is useful to have cheap, manageable and reproducible animal assay systems to test aspects related to bacterial pathogenicity and its control. In this respect, the nematode Caenorhabditis elegans has emerged as a valid animal model. The C. elegans genome sequence indicates that at least 36% of 19.000 predicted proteins have a similar counterpart in humans (Sequencing Consortium, 1998).Thus, in spite of the vast evolutionary distance between nematodes and vertebrates, C. elegans is a valid model system to study bacterial pathogens affecting humans and other hosts. In fact, several bacterial pathogens that cause disease in humans are also able to infect and kill C. elegans (Couillault and Ewbank, 2002). Therefore, this nematode has been used to elucidate virulence mechanisms in Pseudomonas aeruginosa (Gallagher and Manoil, 2001), Burkholderia paseudomallei (O’Quinn et al. 2001) and Salmonella typhimurium (Aballay et al. 2000; Aballay and Ausubel, 2001). These last authors observed, in addition, that S. enteritidis is also able to infect and kill C. elegans. S. enteritidis causes gastrointestinal disease in humans and is a major public health concern due to its ability to be transmitted via contaminated eggs or egg-based and poultry meat products (Dominguez et al. 2002). An approach to the control of S. enteritidis is the use of bacteriophage wich have proven of value in the curtailment S. enteritidis infection in Cheddar cheese (Modi et al. 2001) and vegetables (Leverentz et al. 2001). In this latter context, we have recently isolated and described three dsDNA phages that lyse S. enteritidis in vitro; in addition, variants of these phages, that lyse S. pullorum, can be recovered as well (Santander and Robeson, 2002). These phage have a morphology similar to that of bacteriophage l of Escherichia coli. Furthermore they form clear plaques in lawns of the aforesaid bacteria. Being phage f3aSE the most active we decided to test its potential use in prophylaxis, developing an assay system with C. elegans. C. elegans, as stated above, has been a valuable animal system to test bacterial pathogens with the advantages of ease of manipulation and thorough knowledge of its genetics and molecular biology. Using C. elegans as an assay system we found that the phage f3aSE protects C. elegans from infection and subsequent death by S. enteritidis. We also report that S. pullorum is able to infect and kill C. elegans and that f3aSP, a variant of f3aSE adapted to S. pullorum, protects C. elegans from S. pullorum killing as well. METHODS Bacteria and bacteriophages Bacteria were grown in Luria Bertani medium (LB) at 37ºC. When necessary, the antibiotics (Sigma, St. Louis) rifampicin (Rif; 100 mg/ml) or streptomycin (Sm; 80 mg/ml) were added. A Rif resistant mutant of S. enteritidis ATCC (American Type Culture Collection) 13076 and a strain of S. pullorum (Instituto de Salud Pública-ISP-Chile) were used in infection and phage prophylaxis assays and to propagate phage. Escherichia coli OP50 Sm resistant (Brenner, 1974) was used to feed C. elegans. In addition, we used a strain of S. enteritidis PT4 (ISP, Chile) and S. typhimurium SL1344 (Aballay et al. 2000), the latter as a positive control for pathogen mediated death of C. elegans. The phage used were f3aSE, active against S. enteritidis, and the derivative f3aSP, adapted to lyse S. pullorum (Santander and Robeson, 2002). Liquid lysates (109-1010 pfu/ml) were prepared in LB, with aeration at 37ºC (Santander and Robeson, 2002). Phage were then washed thrice with 0.5 ml of distilled water using a 100 kD filtration unit (Eppendorf centrifugal filter tubes) and finally suspended in 0.5 ml. The suspension was titrated and kept at 4ºC. The nematode C. elegans Bristol N2 was maintained in modified NG agar (0.35% bacto peptone), fed with 200 ml of a fresh culture of E. coli OP50 per plate according to Aballay et al. 2000. For phage prophylaxis assays nematodes were fed, additionally, with 100 ml of phage suspension (108 -109 pfu/ml). We used the method described by Aballay et al. 2000, without transfers. Nematodes were incubated at 25ºC for 10 days. Dead nematode counts were performed every 24 hrs eliminating dead specimens from the plate. Thus, we determined the TD50 (Time it takes for 50% of the nematodes to die). Presence of bacteriophage inside C. elegans Nematodes fed with bacteriophage and E. coli OP50 were suspended in 1.5 ml of S-Basal medium (Brenner, 1974) and washed 20 times by centrifugation (10.000 r.p.m, 5 min.) in a Eppendorf 5415D centrifuge, using 1,5 ml of S-Basal per wash. The last supernatant was titrated for bacteriophage, as well as the nematode pellet after mechanical disruption with a glass pestle fitting 1.5 ml Eppendorf tubes. A control tube treated the same way, except for the mechanical disruption procedure, was used to determine total nematode counts. Bacteriophage prophylaxis assay in C. elegans E. coli OP50 and bacteriophage fed nematodes kept on NG agar plates were used; 15-20 nematodes (L4 stage) were inoculated onto PGS agar plates (Aballay et al. 2000) containing pathogenic Salmonella bacteria. Plates were incubated at 25ºC and dead nematode counts were performed every 24 hrs. We used one way ANOVA and Tukey`s analysis (Sokal and Rohlf, 1979) with an error factor (a) of 0.5% using a Microsoft Excel sheet. Data from the fourth to the tenth day were used (Figure 1 and Figure 2). RESULTS AND DISCUSSIONWe determined that S. enteritidis ATCC 13076 Rif resistant, S. enteritidis PT4 and S. pullorum kill C. elegans as does S. typhimurium SL1344 (Figure 1). The estimated TD50 is the same for both strains of S. enteritidis (S. enteritidis ATCC 13076, 7.02 +/- 0.72 days; S. enteritidis PT4 7.07 +/- 0.12 days), which were more virulent than S. pullorum (7.30 +/- 0.26 days). Both S. enteritidis and S. pullorum were less virulent than S. typhimurium SL1344 (6.40 +/- 0.25 days). These assays were performed without transfers and both eggs and larvae were eliminated, when possible, for the case of the E. coli OP50 grown nematodes. This was not necessary in assays involving S. enteritidis and S. pullorum since no eggs or larvae were observed. It seems that these bacteria interfere with gonad function in C. elegans, in a similar fashion as S. typhimurium does (Aballay and Ausubel, 2001). In all Salmonella infection treatments, the death of C. elegans started at 96 hrs post-infection (Figure 1). The effects of infection were quite evident upon observation of the intestinal tract, which was dilated. This is a typical symptom shown by C. elegans infected by other pathogenic salmonellae, namely S. typhimurium (Aballay et al. 2000). Statistical analyses showed significant differences between infection treatments and the control group of nematodes fed with E. coli OP50. No significant differences were observed with respect to the positive control of C. elegans infected with S. typhimurium SL1344. In relation to the use of bacteriophage in conjunction with C. elegans we first determined that when the nematode was fed E. coli OP50 together with bacteriophage, the latter remained inside the nematode at least during 15 days as determined by their ability to form plaques (pfu) in the corresponding indicator bacteria. For bacteriophage f3aSE we determined that at 15 days of incubation each nematode contains, inside, approximately 21 pfu with an initial input of 109 pfu per culture plate, with contains about 104 nematodes at the time of bacteriophage quantitation. In the case of f3aSP, each nematode contained, inside, about 5 pfu with initial input of 108 pfu/plate. It seems that the bacteriophage are innocuous for C. elegans since we could not observe any abnormalities in nematodes to which bacteriophage were added. Furthermore we found that bacteriophage f3aSE and f3aSP protect C. elegans from killing by S. enteritidis ATCC 13076 and S. pullorum, respectively. When the nematodes containing bacteriophage are exposed to lawns of the bacterial pathogens, they showed significantly less mortality than untreated controls (Figure 2A-B). In addition, no effects were observed in the intestinal tract of bacteriophage protected nematodes. These results suggest that phages such as f3aSE could be used prophylactically in chicken as an additional control measure to diminish S. enteritidis contamination. In summary we have shown that C. elegans can be used in assays of phage prophylaxis in connection with bacterial pathogens able to kill this nematode. ACKNOWLEDGEMENTS We thank Dr. Alejandro Aballay (Harvard Medical School) for the generous gift of C. elegans, S. typhimurium SL1344 and E. coli OP50 and Dra. Soledad Prat, ISP, Chile, for providing additional bacterial strains. REFERENCES Note: Electronic Journal of Biotechnology is not responsible if on-line references cited on manuscripts are not available any more after the date of publication. Supported by UNESCO / MIRCEN network Copyright 2004 by Universidad Católica de Valparaíso -- Chile The following images related to this document are available:Photo images[ej04020f2.jpg] [ej04020f1.jpg] |
| |||||||||

{kind=link}
{kind=link}