 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Electronic Journal of Biotechnology, Vol. 7, No. 2, August, 2004 SHORT COMMUNICATION Efficient identification of tetR-expressing cell lines for tetracycline-regulated gene expression Ping Song1, Hwa-Chain Robert Wang*2 1Department of Pathobiology,
College of Veterinary Medicine,
The University of Tennessee,
2407 River Drive, Knoxville
TN 37996, USA
Tel: 865-974-5837
Fax: 865-974-5616
E-mail: psong3@utk.edu Financial support: National Institutes of Health, USA. Received December 12, 2003 / Code Number: ej04021 ABSTRACT The technology of tetracycline-inducible gene expression has been successfully used in experimental biology to identify the function and downstream signaling pathways of an interested gene. It has been significantly improved to meet the criteria with specificity to exogenous non-toxic inducers, independent regulation from cellular pathways, and dose-dependent inducibility and reversibility. However, to establish tetracycline-inducible gene expression in mammalian cells is still a time-and effort-consuming process. With a tetracycline-inducible gene expression system T-REx, we have developed a practical protocol to use the oncogenic H-ras gene as a dominant reporter gene to increase efficiency in attaining desired cell lines in which ectopic expression of a particular gene in cells can be introduced and reversibly induced by the presence or absence of tetracycline in cultures. Keywords: 10T1/2 cells, Ras, Tetracycline-inducible expression system, tetR, T-REx. Abbreviations: ARTICLE
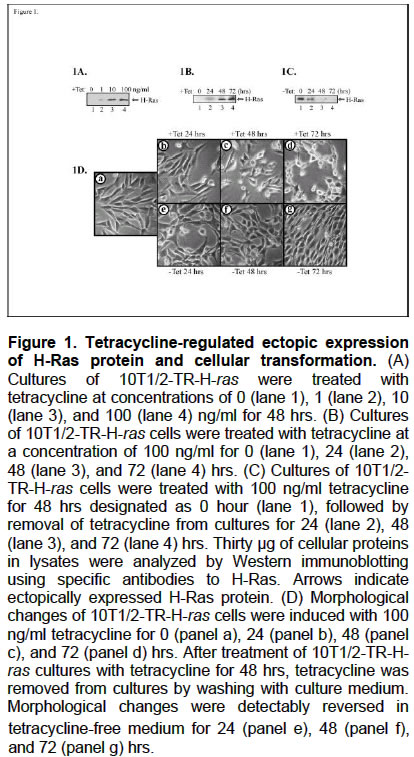
Ectopic expression of gene products in cells is a widely used method in experimental biology to identify the function and downstream signaling pathways of a gene of interest. Using inducible gene expression systems eliminates discrepancy due to non-homogenously transfected cell population with a transient gene expression system and eliminates inconsistency due to clonal variation with a stable gene expression system (Rossi and Blau, 1998; Tate et al. 2003; Xu et al. 2003). The tetracycline-inducible expression system uses non-mammalian control elements that do not have the ability to respond to cellular factors, to interfere with cellular pathways, or to bind genomic DNA in mammalian cells (Gossen and Bujard, 1992; Gossen et al. 1995; Tate et al. 2003). In the past ten years, tetracycline-inducible gene expression systems have been significantly improved to meet the criteria for an ideal system with specificity to exogenous non-toxic inducers, independent regulation from cellular pathways, and dose-dependent inducibility and reversibility. However, to establish tetracycline-inducible gene expression in mammalian cells is a time-and effort-consuming process. There is urgent need of a practical protocol to efficiently construct a cell line in which ectopic expression of a specific gene is controlled by a tetracycline-inducible gene expression system. We have used a tetracycline-inducible T-REx expression system which consists of two key expression vectors pcDNA4/TO-E and pcDNA6/TR (Yao et al. 1998). The pcDNA4/TO-E vector contains complete cytomegalovirus enhancer-promoter (PCMV) and two tetracycline operator sites (tetO) for PCMV/tetO-regulated gene expression in mammalian cells. In the presence of the tetracycline repressor (TetR) that is expressed by pcDNA6/TR, the tetO sites in pcDNA4/TO-E are bound with TetR and transcription is repressed. In the presence of tetracycline, tetracycline binds to TetR and releases TetR from the tetO sites, allowing transcription to proceed. Accordingly, increasing efficiency in identifying and isolating a cell clone which expresses adequate levels of TetR to effectively control PCMV/tetO-regulated gene expression is the key step to the construction of a cell line in which ectopic expression of a specific gene is tightly controlled by this tetracycline-inducible gene expression system. In this communication, we demonstrated that using the oncogenic H-ras gene (Campbell et al. 1998) as a dominant reporter gene helping in effectively identifying pcDNA6/TR-transfected cell clone which is competent to support PCMV/tetO-regulated gene expression. MATERIALS AND METHODS Mouse embryo fibroblast 10T1/2 cells in a 35 mm culture dish were transfected with 2 μg of pcDNA6/TR plasmid DNA (Invitrogen, Carlsbad, CA, USA) with PolyFect Transfection Reagent (Qiagen, Valencia, CA, USA). 10T1/2 cells were routinely maintained in Basal Medium Eagle (BME) (Life Technologies, Grand Island, NY, USA) supplemented with 10% fetal bovine serum, 100 units/ml penicillin, and 100 μg/ml streptomycin, and incubated at 37ºC (Fecteau et al. 2002). Cells were routinely subcultured every 3 or 4 days. After 48 hrs of transfection, cells were subcultured into three 100 mm culture dishes. Transfected cultures were treated with a selective agent blasticidin (Invitrogen) at 10 μg/ml. The oncogenic H-ras gene was used as a dominant reporter gene and constructed into pcDNA4/TO-E vector (Invitrogen), resulting in pcDNA4/TO-E-H-ras. 10T1/2 cell lines which were stably transfected with pcDNA6/TR in 35 mm culture dishes were super-transfected with 1.5 μg of pcDNA4/TO-E-H-ras plasmid DNA. After 48 hrs of transfection, cells were subcultured into three 100 mm culture dishes. In addition to 10 μg/ml blasticidin, Zeocin (Invitrogen) was added into cultures at 200 μg/ml to select cell clones which had been stably transfected with both pcDNA6/TR and pcDNA4/TO-E-H-ras. The Krs1 gene (Taylor et al. 1996) that does not exhibit a dominant phenotype and was fused with a tag peptide Flag (Knappik and Pluckthun, 1994) at its N-termini was constructed into pcDNA4/TO-E vector, resulting in pcDNA4/TO-E-Flag-Krs1. The 10T1/2-TR cell line was super-transfected with 1.5 μg of pcDNA4/TO-E-Flag-Krs1 plasmid DNA and incubated with both 10 μg/ml blasticidin and 200 μg/ml Zeocin to select cell clones which had been stably transfected with both pcDNA6/TR and pcDNA4/TO-E-Flag-Krs1. In addition, pcDNA4/TO-E-H-ras and pcDNA4/TO-E-Flag-Krs1 were individually super-transfected into cultures of T-REx293 cells (Invitrogen), which are stably transfected with pcDNA6/TR. Super-transfected cultures were incubated with both 5 μg/ml blasticidin and 400 μg/ml Zeocin to select cell clones which had been stably transfected with both pcDNA6/TR and pcDNA4/TO-E-H-ras or pcDNA4/TO-E-Flag-Krs1. T-REx293 cells were routinely maintained in Dulbecco’s modified Eagle’s medium (DMEM) (Life Technologies) supplemented with 5% fetal bovine serum, 100 units/ml penicillin, and 100 μg/ml streptomycin, and incubated at 37ºC. T-REx293 cultures were routinely subcultured every 2 to 3 days. To detect ectopic expression of H-Ras and Flag-Krs1, cell pellets were incubated in lysis buffer (10 mM Tris-HCl, 150 mM NaCl, 1% Triton X-100, 5 mM EDTA, 10 mM sodium pyrophosphate, 10% glycerol, 0.1% Na3VO4, 50 mM NaF, pH 7.4) on ice for 10 min (Fecteau et al. 2002). Cell lysates were isolated from the supernatants after centrifugation of crude lysates at 20,000 x g for 20 min. Protein concentration in cell lysates was measured using the BCA assay (Pierce, Rockford, IL, USA). Protein levels of H-Ras and Flag-Krs1 in cell lysates were detected by Western immunoblotting using a Ras-specific antibody (Santa Cruz Biotechnology, CA, USA) and Flag-specific antibody (Sigma), respectively, with the Supersignal chemiluminescence kit manufacturer (Pierce), visualized by autoradiography. RESULTS AND DISCUSSION Seven days after transfection of 10T1/2 cells with pcDNA6/TR plasmid DNA, 8 cell colonies were cloned and developed into cell lines. According to typical 10T1/2 cell morphology, 5 cell lines were chosen for the identification of cell lines which are capable of supporting PCMV/tetO-regulated gene expression. After super-transfection of these cultures with pcDNA4/TO-E-H-ras plasmid DNA for 7 days, we identified 42 cell colonies which exhibited normal morphology of 10T1/2 cells and 59 cell colonies which exhibited ras-transformed morphology in one of the five pcDNA6/TR-transfected 10T1/2 cell lines. After addition of 100 ng/ml tetracycline into cultures of this line of cells for 48 hrs, 24 out of 42 cell colonies with normal morphology of 10T1/2 cells came to exhibit distinct ras-transformed rounded refractile morphologies. We designed this cell line as 10T1/2-TR. In comparison, colonies formed in cultures of the other 4 cell lines did not show distinct changes between normal and ras-transformed morphologies in the presence or absence of tetracycline in their cultures. Cell colonies which were induced to exhibit transformed morphology distinctly by tetracycline were selected and established as cell lines, designated as 10T1/2-TR-H-ras. Addition of tetracycline into cultures of 10T1/2-TR-H-ras cells induced expression of H-Ras protein to various levels in a dose-dependent manner (Figure 1A) and induced progressive increases of ectopically expressed H-Ras protein in a time-dependent manner (Figure 1B). Removal of tetracycline from the transformed 10T1/2-TR-H-ras culture resulted in progressive decreases of ectopically expressed H-Ras protein (Figure 1C). Concomitantly, transformation of cell morphology was induced in 10T1/2-TR-H-ras cultures from normal spindle morphology (Figure 1D, panel a) to typical rounded refractile ras-transformed morphology (panels b-d). Depletion of tetracycline from transformed cultures reversed cell morphology to normal spindle morphology (Figure 1D, panels e-g). Ectopic expression of H-Ras protein and cellular transformation of 10T1/2-TR-H-ras cells are reversibly controlled by the presence or absence of tetracycline in cultures in a time- and dose-dependent manner. To test the applicability of the identified 10T1/2-TR cell line to support PCMV/tetO-regulated expression of other transgenes which do not exhibit a dominant phenotype, the identified 10T1/2-TR cell line was super-transfected with pcDNA4/TO-E-Flag-Krs1 plasmid DNA. After selection with both blasticidin and Zeocin (Invitrogen), tetracycline-induced ectopic expression of Flag-Krs1 (Figure 2A) was detected in 2 of 10 arbitrarily isolated cell colonies of 10T1/2-TR-derived cells. Using the commercially available tetR-expressing cell line T-REx293 (Invitrogen) as a comparison counterpart cell line, tetracycline-induced ectopic expression of Ras and morphological changes were detected in approximately 30% of transfected cell colonies of T-REx293 cells. Tetracycline-induced ectopic expression of Flag-Krs1 (Figure 2B) was detected in 2 of 5 arbitrarily isolated cell colonies of T-REx293 -derived cells. Apparently, the identified 10T1/2-TR cell line is competent to support PCMV/tetO-regulated gene expression and the T-REx system is adequate to establish tetracycline-induced expression cell lines. In addition, we did not detect any discrepancy of results between using tetracycline-free fetal bovine serum (Clontech, Palo Alto, CA) and qualified fetal bovine serum (Life Technologies) in culture medium in this study. In conclusion, the method reported in this communication provides a practical protocol to develop a desirable cell line in which ectopic expression of a gene of interest is reversibly induced by the presence or absence of tetracycline in cultures. Effectively isolating a competent tetR-expressed cell line is the key step to increase the efficiency of attaining a desired cell line in which ectopic expression of a specific gene can be reversibly regulated by a PCMV/tetO system. Conventionally, at least 20 lines of pcDNA6/TR-transfected cells are selected to test for tetracycline-inducible gene expression by transiently transfecting with the positive control plasmid expressing β-galactosidase. Biochemical assays are required to detect tetracycline-induced expression of β-galactosidase. In our study, tetracycline-induced ectopic expression of H-Ras protein is phenotypically and responsively revealed by morphological transformation of cells that can be initially monitored microscopically. Subsequently, ectopically expressed protein can be verified by biochemical assays. Using the transforming H-ras gene as a reporter gene shows a clear advantage to efficiently identify cell clones in which TetR is adequately expressed to tightly control PCMV/tetO-regulated gene expression. ACKNOWLEDGEMENTS We thank Dr. K. A. Fecteau for critical review of this manuscript. REFERENCES
Note: Electronic Journal of Biotechnology is not responsible if on-line references cited on manuscripts are not available any more after the date of publication. Supported by UNESCO / MIRCEN network Copyright 2004 by Universidad Católica de Valparaíso -- Chile The following images related to this document are available:Photo images[ej04021f1.jpg] [ej04021f2.jpg] |
| |||||||||

{kind=link}
{kind=link}