 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Electronic Journal of Biotechnology, Vol. 7, No.3, December, 2004, pp. 189-190 RESEARCH ARTICLE Controlling proteolytic degradation of the methionine enriched MB-1Trp protein Maxime Sasseville1,Catherine St-Louis2,Habib
Khoudi3,Marc Beauregard*4 *Corresponding author Financial support: This
work was supported by grants from CORPAQ (Gouvernement du Québec) and NSERC
( Code Number: ej04026 ABSTRACT Protein design is currently used for the creation of new proteins with desirable traits, which include a superior nutritional value. One of the challenges of protein design in this area is to achieve the production of stable native-like proteins that resist the proteolytic pressure of the organism used for its production (the bioreactor). We report here the identification of a specific peptide bond sensitive to E. coli proteolysis in the designer protein MB-1Trp. In an attempt to reduce proteolysis, we have created a MB-1TrpHis gene library in which the two amino acids surrounding the peptide bond, N44 and L45, were randomized using degenerated oligonucleotides. The initial characterization of MB-1TrpHis N44E/L45V and MB-1TrpHis N44E/L45M, 2 variants of the library that were more resistant than the parent protein, was performed in order to investigate the nature of the mutants’ resistance. Our results suggest that the mutants behaved like MB-1Trp regarding folding and thermal stability, and that proteolytic resistance is due to the elimination of the protease recognition site. Keywords: agro-biotechnology, essential amino acids, protein design, protein engineering, proteolysis. Abbreviations: ARTICLE The control of proteolysis is crucial in a number of protein applications and in several cellular events (Pipe and Kaufman, 1997; Morrow et al. 2002). In agro-biotechnology, BT toxin which is expressed in insect resistant crops has been engineered to resist a specific inactivating proteolysis by insect gut proteases (Audtho et al. 1999). Similarly, Wyss and coworkers have engineered a longer half-life phytase by decreasing its proteolytic susceptibility, allowing a better assimilation of phytic acid by monogastric farm animals (Wyss et al.1999). The expenses related to feed additives used for animal production has led to the study of intracellular production of high-quality proteins by transgenic crops and other organisms as a means of obtaining efficient and less costly sources of essential amino acids (EAA) (Dyer et al. 1993; Ealing et al. 1994; Williamson et al. 1996; Forano et al. 2000). Previous attempts to express methionine-rich proteins in different crops achieved various levels of success (Kirihara et al. 1988; Guerche et al. 1990; Altenbach et al. 1992; Aragao et al. 1992; Saalbach et al. 1994; Chaudhuri et al. 1995; Dyer et al. 1995; Teuber et al.1998; Alcocer et al. 2002). Interesting results have been reported in the manipulation of EAA contents through metabolic engineering (Galili and Hofgen, 2002). Approaches that combine both strategies (protein production and metabolic engineering) have been reported in recent patents (Falco et al. 1996; Beach and Tarczynski, 2000). The issue of proteolytic stability is also of paramount importance for the production of such high value proteins. In this regard, de novo design of proteins offers a powerful way to deliver high quality nutrients for animal nutrition, but earlier attempts lead to proteins that were marginally stable and/or had a poor behaviour in vivo (Doel et al. 1980; Jaynes et al. 1985). In the early 1990s, we met the stability challenge inherent to designer proteins and created a de novo designed sink protein, MB-1, that was shown to have a folded core and a low affinity for 8-anilino-1-naphthalenesulfonic acid (ANSA) (Beauregard et al. 1995). Its behaviour and expression levels in vivo were found to be far superior to those obtained from earlier attempts in this field of research (MacCallum et al. 1997). MB-1 was demonstrated to work efficiently as a sink for EAA, with an important advantage over its natural counterpart: the controlled proportion of each EAA included in its sequence. Despite this initial success, the designer protein MB-1 had a low conformational stability and was only marginally resistant to proteases at physiological temperature (McCallum et al 1997). Such results suggest that MB-1 would not accumulate in bioreactors such as transgenic organisms, where it would be exposed to endogenous proteases (Liao, 1993). A second
generation mutant, named MB-1Trp, was generated and was significantly more
thermoresistant than its parent protein MB-1, by up to In order to improve MB-1Trp resistance to proteolytic degradation, the localization and the modification of sensitive residues was undertaken. Using a combinatorial approach, the residues flanking the targeted bond were substituted by similar residues based on their predicted positions in the protein. Two stabilized mutants of the library were further characterised in vitro in order to understand the effect of the mutations on MB-1Trp stability and behaviour.
MATERIALS AND METHODS
MB-1Trp expression and purification MB-1Trp
was expressed and purified according to the method described previously (Gagnon
et al. 2000). Protein samples intended for mass spectrometry were purified
on a DEAE-Sepharose column, equilibrated with Matrix Assisted Laser Desorption Ionisation-Time of Flight (MALDI-TOF) mass spectrometry The size of the fragments was determined using an Applied Biosystems MALDI-TOF Mass Spectrometer Voyager System 2016 with an a-cyano-4-hydroxycinnamic acid matrix. WinPep software (Hennig, 1999) was used to calculate the mass of the fragments as deduced from sequence. MB-1TrpHis construction, expression and purification To allow a C-terminal specific purification of MB-1Trp and its fragments, and detection with a commercially available anti-His-Tag monoclonal antibody, a poly-histidine tag (His-tag) was ligated to the C-terminal end of MB-1Trp. This was achieved by constructing a chimeric plasmid from the original pMal-c2 MB-1Trp and pMal-c2 MB-1His plasmids (Grundy et al. 1998). Protein samples for N-terminal sequencing were purified by amylose affinity chromatography followed by an immobilized metal affinity chromatography (IMAC), as described earlier (Grundy et al. 1998). Proteins
were transferred to a polyvinylidene fluoride (PVDF) membrane as described
by Geisow and Aitken (1989). The proteins were then sequenced
using an ABI model 473A Protein Sequencer (Service Protéomique de l’Est du
Québec, Preparation of MB-1TrpHis mutants Substitutions
of asparagine in position 44 and leucine in position 45 of MB-1TrpHis were
performed using the "Site-Directed Mutagenesis Kit" (Stratagene).
The asparagine codon (AAC) was replaced by a GA(A/T) codon, encoding for
E or D. The leucine codon (CTG) was replaced by a (A/T/G)T(G/C) codon, encoding
V, M, I, L and F in the following proportion: 2:1:1:1:1 respectively. A total
of ten different mutation sets were therefore available with this strategy.
The mutagenic oligonucleotides, purchased from Biocorp
(Montréal, MB-1TrpHis: 5’-CAATG AAG AAT CAT CTT CAA AAC CTG ATG CAG AAG ACT AAG AAC-3' Oligo NL#1: 5’-CAATG AAG AAT CAT CTT CAA GAW DTS ATG CAG AAG ACT AAG AAC-3' Oligo NL#1-R: 5’-GTT CTT AGT CTT CTG CAT SAH WTC TTG AAG ATG ATT CTT CATTG-3' where W stand for A or T; D stand for A, T or G; S stand for C or G; and H stand for A, C, or T. The mutations
were confirmed by dideoxynucleotide DNA sequencing. The mutated genes were
expressed in fusion with the Maltose-Binding Protein (MBP) in pMal-c2 expression
vectors ( Proteolytic resistance screening by western blotting E.
coli XL-1Blue cells (recA1, endA1, gyrA96, thi-1, hsdR17(rK-,mK+), supE44, relA1, lac-,
[F' proAB, lacIqZΔ M15, Tn10(Tetr)])
(Stratagene, Cedar Creek,
USA) harbouring pMal-c2 MB-1TrpHis mutant plasmids were grown in 15
mL of LB Miller medium (EM
Science, Hawthorne, USA) supplemented by 100 mg l-1 ampicillin (Sigma)
to an optical density of 0.3 at 600 nm. Transcription was induced by adding Following
incubation with E. coli proteases, the solutions were denatured using
2% SDS and heated at Expression and purification of MB-1TrpHis mutants All mutants were expressed and purified as described in Gagnon and coworkers (Gagnonet al. 2000) with a second chromatography involving metal affinity purification using the modifications cited above. Protein quantification and purity evaluation For all experiments, protein concentration was determined by bicinchoninic acid (BCA) assay (Sigma) using bovine serum albumin as a standard. SDS-PAGE experiments were conducted prior to measurements to confirm protein purity. Conformational investigation by Circular Dichroism (CD) CD measurements
were carried out as described by Gagnon and coworkers (Gagnon
et al. 2000). Experiments were conducted at a protein concentration of
0.6 mg ml- The samples
were prepared as described in the previous section and conformational changes
were monitored by CD. To measure thermostability, temperature was increased
from 5 to
RESULTS
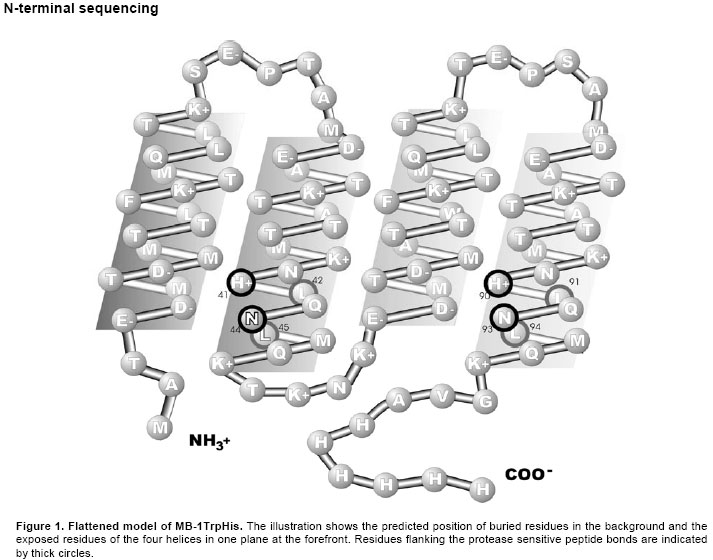
Identification of the protease sensitive peptide bonds Mass spectrometry was performed on MB-1Trp fragments purified using anionic exchange purification (as detailed in Grundy et al. 1988). The assignment of mass spectrometry results is described in Table 1. Seven fragments were tentatively assigned, the ones with the largest signal being M1-N93 (peak no. 1) and M1-N44 (peak no. 3). For some fragments slight differences in calculated and measured masses were found. They might be ascribed to mass changes caused by ionisation and/or post-translational modifications, such as methionine oxidation, which is not unexpected considering the high methionine contents of MB-1Trp (Fontana et al. 1997; Hollemeyer et al. 2002). The assignment of peaks no. 1 and 3 was confirmed by N-terminal sequencing. To facilitate the fragment identification process, N-terminal sequencing was performed on MB-1 TrpHis fragments recovered from an IMAC column. Only fragments that carry the intact C-terminus are retained under such conditions. After recovery from E. coli, several fragments were obtained, the two most important bearing the sequence: LMQKTKNKE and LMQKGVAH at their N-terminus respectively. Such sequences correspond to amino acids 45-53 and 94-101 respectively. Therefore, both approaches (mass spectrometry and N-terminal sequencing) point to a cleavage between N44 and L45, and a second site at N93 and L94. The detection of the second site is not unexpected, because in MB-1Trp, helices 2 and 4 are predicted equivalent regarding both conformation and sequence (per design, Figure 1). Furthermore, the correlation between fragments size and N-terminal sequencing results suggest that adding the His tag to MB-1Trp did not change its sensitivity to proteases.
Another minor fragment had the amino-terminus sequence LQNLMQ, which corresponds to either segment 42-47 associated to peak no. 5 (L42 – A 100) or 91-96, associated to peak no. 2 and 6 (M1-H90 and L45-H90 respectively). The population of fragment that has been cut using the site N44-L45 (Fragment M1-N44, peak no. 3) appeared more important than the ones being cut at the site H41-L42 (Fragment M1-H41 peak no. 4 and fragment L42-A100, peak no. 5). Similarly, the fragment population of M1-N93 (peak no. 1) was more important than the M1-H90 population (peak no. 2). Thus, the N-L bonds appear to be more susceptible or accessible than H-L bonds, probably because they are more exposed as predicted per design (Figure 1). Among the two N-L bonds identified, the N44-L45 bond was selected for our study. A cleavage right in the middle of the protein was believed to have a more important impact on protein integrity and “residual” resistance after a first cleavage. The predicted positions of the two amino acids targeted by proteolysis (N44, L45) are shown in Figure 1. It was hypothesised that changing the residues directly involved at this peptide bond would decrease sensibility to a putative specific protease of E. coli. The N44 residue was allowed to be mutated to D or E, two acidic residues. Interactions with the neighbouring basic residues, H41 and K48, could promote the formation of stabilizing intrahelical salt-bridges. The structural stabilisation of this region of helix 2 could also contribute to prevent proteolytic activity. The L45 residue was allowed to be mutated to the hydrophobic amino acids, I, V, M, F and the wild-type L. With this strategy, we hoped that the most accommodating residues would result in more stable mutants while providing a change in the amino acid sequence at the target site. Since L45 is predicted to be buried in the hydrophobic core, where mutations are usually not well tolerated, a wide variety of non-polar amino acids was allowed (Sanchez-Ruiz and Makhatadze, 2001). Clones generated by the mutagenesis reaction were sequenced to assess the distribution of the mutations in our bank. Seven out of 10 possible combinations were recovered from the mutant library as shown in Table 2. Interestingly, both possible mutant with a conserved L in position 45 were absent from the clones sampled for sequencing. There was no obvious bias in the distribution of the mutations.
Screening mutants for proteolytic resistance Randomly
selected mutants were screened for their proteolytic resistance by exposing
them to proteolytic degradation assays, followed by an analysis by Western
blotting with an anti-His-Tag monoclonal antibody. For degradation assays,
MB-1TrpHis (containing a His tag) was used as the "wild type" protein
instead of MB-1Trp. Proteins were exposed to bacterial proteases before and
after the proteins were separated by factor Xa from their fusion partner
MBP. As can be seen on Figure 2, several mutants outperformed
their parental counterpart MB-1TrpHis when exposed to E. coli proteases:
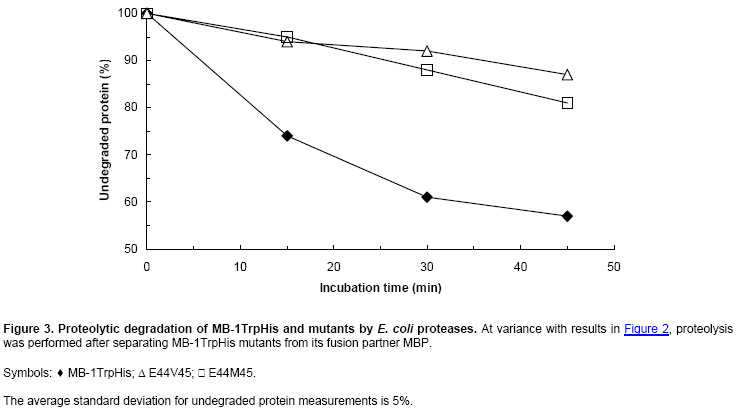
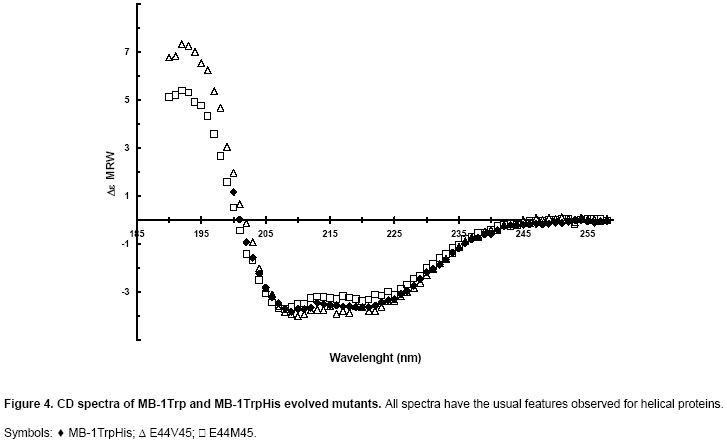
The disappearance of MB-1TrpHis after a 30 min incubation at Careful examination of Figure 2 allows the detection of several protein bands that differ from MB-1TrpHis and its mutants (band position shown by an arrow). In the upper part of the gels, the first band at the top is the MBP-MB-1TrpHis fusion protein that escaped cleavage with factor Xa, migrating at 54 kDa. The next band is the large cleavage product of the fusion: MBP, running at 43 kDa. Then, few bands in the mid-range (25-35 kDa) are believed to be E. coli proteins with histidines at their surface, resulting in some affinity for the antibody used in this experiment. The screened clones were sequenced to assess the distribution of the stabilizing mutations. The most stabilized mutants carry the mutation combinations E-V, E-M, D-I, D-V and D-M. The least stable mutants carry the mutation combinations E-F (lanes 16-17) and D-F (lanes 9-10). These results could be explained by the fact that the hydrophobic core might not be able to harbour the large phenylalanine side-chain in position 45, which could promote local unfolding of the protein and expose protease targets (Goldberg et al. 1978; Betz et al. 1993). The impact of the fusion partner on the mutant resistance to proteolysis was investigated in Figure 3. For these results, the fusion proteins MBP-MB-1Trp mutants were cleaved prior to incubation with bacterial proteases. Figure 3 shows a comparison of MB-1TrpHis with the most stable mutants (MB-1TrpHis E44V45, referred to as “E44V45”, and MB-1TrpHis E44M45, renamed “E44M45”). While 40% of MB-1TrpHis has disappeared after 40 min incubation with proteases, only 10-15% of both mutants initial populations was lost to proteolysis. Such results confirm the intrinsic stabilisation of the nutritive protein MB-1TrpHis by the mutations performed here and the absence of interference due to some protective effect from the fusion partner MBP. Note that Figure 2 was generated using antibody to the His tag, while Figure 3 describes results obtained using antibody to MB-1Trp. Therefore, the specificity of the detection protocol has no impact on the apparent stability of the designer proteins under study. Smaller protein fragments (probably MB-1TrpHis proteolytic products) were detected in the parent protein electrophoretic pattern and for most mutants. They appear to be less important for the mutants however, when considering the relative amount of full length product vs degradation bands. Effect of mutations on secondary structure Secondary structures of the two most stable mutants MB-1TrpHis N44E/L45V and MB-1TrpHis N44E/L45M were analysed by CD. The spectra measured for the mutants were typical of helical proteins and equivalent to MB-1Trp (Gagnon et al. 2000) as shown in Figure 4. The two mutants’ secondary structures were calculated using CDSSTR algorithm (data not shown). No significant changes in the secondary structures were detected, which indicates that proteolytic resistance is not the consequence of conformational changes that would promote hindrance for protease access to its target. Effect of mutations on conformational stability Thermal
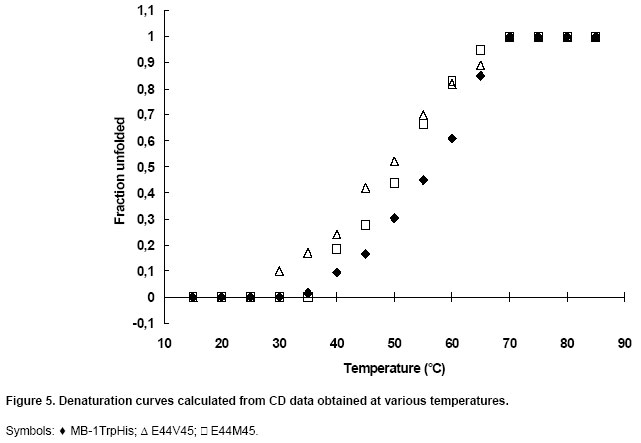
denaturation experiments were carried out on the two purified mutants using
CD as described above in Materials and Methods. Their
melting temperatures (Tm) are
DISCUSSION
The design of novel proteins with native-like properties is, thus far, still a challenging issue. Many early attempts have given ambiguous results demonstrating molten globule states, loosely packed proteins and polydisperse protein populations (Hill et al. 2000). In particular, b-based structures have proven difficult to design in a highly soluble state or without aggregation problems (Beauregard et al. 1991; Jin et al. 2003). Recent improvements in core modelling and packing algorithm have lead to engineering of well behaved proteins (Dahiyat et al. 1997). It is worth mentioning that such computer generated cores do not include significant amount of Met, if any, and their AA composition is not among the criteria used for their design. Obviously, designer proteins did not go through billions of years of optimisation via evolution like natural proteins did. This in part may explain the poor characteristics of several de novo designed proteins. Another factor is our limited comprehension of all the rules that govern protein folding and dynamics. Nutritious designer proteins are not exempt from such limitations, and early attempts have revealed low stability and low protein yield in vivo (Doel et al. 1980; Jaynes et al. 1985). Instead of modifying their protein of interest, other groups have modified the production host by disabling the specific protease that digest the protein (Heo et al.2002). Obviously, such a strategy cannot be of application for all available bioreactors. Further, the use of nutritive proteins for ruminants requires a high degree of intrinsic resistance to proteases for them to behave as by-pass proteins and have an optimal impact on animal performances (Stern et al.1985). We reported here a continuation of our rational approach to designing stable proteins with the additional constraint represented by a biased content in selected EAA (M, T, K and L). The mutations introduced to the protein MB-1Trp were shown to have an important impact on its resistance to E. coli proteases (Figure 3). This alone is a major improvement in MB-1Trp behaviour. Enhancement of resistance to proteolysis afforded by specific interactions with the fusion partner MBP was considered and ruled out: the evolved proteins studied here were more resistant before and after cleavage from MBP. In order to understand the nature of the stabilization brought by the mutations, their effect on MB-1Trp conformation was assessed. As shown by CD and thermal denaturation experiments, these contributions appeared marginal. This implies that the improvement probably came from the simple deletion of the targeted sequence recognised by the protease(s). The improvement in resistance achieved by modifying residues 44 and 45 supports the validity of our strategy in selecting this particular area as described above. The sites of proteolysis could be informative on the secondary structure locations. As proteases cut in loop or accessible structures preferably (Yoo et al. 2001; Heo et al. 2002), one could speculate that the region around position 44-45 would not be in a helical conformation as predicted, but would rather be in unordered or flexible conformation. This would be in agreement with the helical content of MB-1Trp, which is lower than expected per design (55% vs 75%). Premature ending of the second and fourth helices could lead to exposure of these regions. Further experiments including non-specific proteinase K degradation will help us to locate more precisely the helices, as they are not cleaved under those conditions (in globular protein in native state, short incubation times and low protease concentration; Polverino de Laureto et al. 1995). However, as the targeted bonds were identified to be the same in second and fourth helices, it gives us evidence that the structure around the homologous site Asn93-Leu94 could be quite similar and that our strategy, applied to these two positions, would similarly improve the behaviour of MB-1Trp, at least for the yield of full length protein. Our strategy of protease target location reveals itself to be an effective and simple way to increase MB-1Trp half-life in a normal cellular environment. It would be interesting to see how this set of mutations will affect the accumulation of our protein in plant cytosol for example. Our goal is to improve the stability of EAA enriched proteins to a level comparable to that of natural crop proteins. ACKNOWLEDGMENTSThe authors
are indebt to Dr Catherine Servis at Institut de Biochimie, Université de
Lausanne (
Note: Electronic Journal of Biotechnology is not responsible if on-line references cited on manuscripts are not available any more after the date of publication. Copyright 2004 by Universidad Católica de Valparaíso -- Chile The following images related to this document are available:Photo images[ej04026f3.jpg] [ej04026f1.jpg] [ej04026f4.jpg] [ej04026f2.jpg] [ej04026f5.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}