 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Electronic Journal of Biotechnology, Vol. 7, No.3, December, 2004, pp. 256-263 RESEARCH ARTICLE Influence of various fermentation variables on exo-glucanase production in Cellulomonas flavigena Muhammad Ibrahim Rajoka Code Number: ej04029 ABSTRACT The influence of carbon and nitrogen sources on the production of exo-glucanase was investigated. The enzyme production was variable according to the carbon or nitrogen source used. Levels of b-cellobiohydrolase (CBH) were minimal in the presence of even low concentrations of glucose. Enzyme production was stimulated by other carbohydrates and thus is subject to carbon source control by easily metabolizable sugars. In Dubos medium, on cellobiose, the cellobiohydrolase titres were 2-to 110-fold higher with cells growing on monomeric sugars and 2.7 times higher than cells growing on other disaccharides. a-Cellulose was the most effective inducer of b-cellobiohydrlase and filter paperase (FPase) activities, followed by kallar grass straw. Exogenously supplied glucose inhibited the synthesis of the enzyme in cultures of Cellulomonas flavigena. Nitrates were the best nitrogen sources and supported greater cell mass, cellobiohydrolase and FPase production. During growth on a-cellulose containing 8-fold sodium nitrate concentration, maximum volumetric productivities (Qp) of b-cellobiohydrolase and FPase were 87.5 and 79.5 IU/l./h respectively and are significantly higher than the values reported for some other potent fungi and bacteria. Keywords: Carbohydrates, cellobiohydrolase, Cellulomonas flavigena, exo-glucanase, enthalpy, entropy, fermentation, induction. ARTICLECellulases and xylanases are implicated in baked foods, fruit processing, cloth cleaning, preparation of dehydrated vegetables and food products, preparation of essential oils, flavours, pulp and paper production, starch processing, preparation of botanical extracts, jams, baby foods, juices, degumming coffee extracts, improved oil recovery, waste treatment, textile refining and preparation of feed for farm animals (Kubicek et al. 1993; Hoshino et al. 1997; Lynd et el. 2002). Cellulase
production has attracted a world-wide attention due to the possibility of
using this enzyme complex for conversion of abundantly available renewable
lignocellulosic (LC) biomass for production of carbohydrates for numerous
industrial applications including bioethanol (Gadgil et al.
1995; Hayward et al. 2000). Economical production of
cellulases is key for feasible bioethanol production from LC biomass using
cellulase-based processes. Different fungi and bacteria have been used for
production of cellulases and xylanases using different substrates (Bahkali,
1996; Magnelli and Forchiassin, 1999; Shin
et al. 2000; Lynd et al. 2002). Previously we produced
cellulases following growth of C. biazotea on different substrates
produced on saline lands namely Leptochloa fusca (kallar grass) straw, Panicum
maximum, Sesbania aculeata compared with bagasse straw, and wheat
straw and found that kallar grass was superior to other substrates for supporting
synthesis of cellulases (Rajoka and Malik, 1997). Cellulase
system of C. flavigena which produced high activities of cellulases
and xylanases during growth on kallar grass showed end-product inhibition-resistance
and thermal stability at room temperature of These enzyme characteristics prompted to further investigate the production potential of FPase, cellobiohydrolase or exoglucanase (EC 3.2.1.91) as it measures the complete cellulase complex (Duenas et al. 1995). Celluloses and the cellulosic components in LC substrates are essential for formation of mRNA to support maximum formation of cellulases at the transcription level (Gutierrez-Nova et al. 2003). Glucose, on the other hand, represses cellulase synthesis by a catabolite repression mechanism at the transnational level (Rajoka et al. 1998; Ponce-Noyola, 2001; Lockington et al. 2002). A study of b-cellobiohydrolase, measured on r-nitro-phenyl b-D-cellobiopyranoside and FPase, measured on filter paper (which correlated with substrate utilization parameters) by C. flavigena including its induction, repression, and production is presented. MATERIALS AND METHODSMicro-organism and growth medium Cellulomonas
flavigena NIAB 441 (Rajoka and Malik, 1997) was used
in these studies. Stock cultures of the organism were stored in culture
tubes using Dubos salt-cellulose agar plates after the instruction of the
stock culture maintainers and grown in liquid cultures as described earlier
(Rajoka and Malik, 1997). All chemicals were of analytical
grade. Carboxymethylcellulose (Na-salt) of low viscosity and a-cellulose
were from Sigma Chemical Co., Evaluation of carbon sources for induction of exo-glucanase A total of 12 different carbon sources in triplicate (Table 1) were chosen as test substrates on the basis of literature data and their availability. The organism was grown and enzyme preparations processed as described earlier (Rajoka and Malik, 1997).
Effect of nitrogen sources on cellulase production Different nitrogen sources (Table 2) were tested to select the best nitrogen source keeping nitrogen concentration at 0.164 g/l. The cultures were grown in time course study as described earlier and processed for enzyme assays. Sodium nitrate proved to be the best nitrogen source and its concentration was optimized by changing its concentration in the medium while maintaining other optimal cultural conditions.
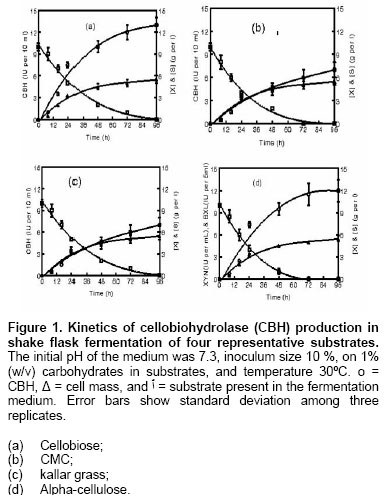
The ability of the organisms to utilize monosaccharides, disaccharides, kallar grass, with reference to cellobiose, CMC, or a-cellulose (Avicel) as sole carbon source was examined in basal Dubos salts medium containing 0.2% yeast extract as described earlier (Rajoka and Malik, 1997). Carbon sources were added individually to batches of basal medium to give a saccharide level of 10 g/l. All monosaccharides and disaccharides were added to autoclaved medium after filter-sterilization. All media were adjusted to pH 7.3 with 1 mol NaOH or 1 mol HCl and were dispensed in 200 ml aliquots into 1-l Erlenmeyer flasks in triplicate. For determining
enzyme synthesis, above medium was inoculated with 10% of inoculum containing For cellulolytic
enzyme assays, the appropriately diluted culture supernatant or cell extract
(Rajoka and Malik, 1997) was used to determine FP cellulase
(FPase) activity or cellobiohydrolase activity using filter paper no.1 or para-nitrophenyl-b-D-cellobioside
(Sigma) respectively in 0.2 mol
acetate buffer (pH 7) at Saccharides were determined using 3,5-dinitrosalicylic acid after Miller (1959). Cellulose and hemicellulose were determined as described previously (Latif et al. 1994). Cellobiosidase
was mainly cell-bound while FPase was secreted in the medium. Therefore,
the proteins in both extracellular and cellular fractions were determined
according to Determination of kinetic parameters Allkinetic parameters for substrate consumption and product formation (Rajoka and Malik, 1997) were determined as described previously (Rajoka et al. 1998). RESULTS AND DICUSSIONExtensive screening of potential cellobiosidase, (CBH) or filter paperase (FPase) inducers showed that when Cellulomonas flavigena NIAB 441was cultured on media containing monomeric saccharides, dimeric saccarides, carboxy methyl cellulose (CMC), a-cellulose or kallar grass (Table 1), it had a shorter lag period and more specific growth rate when grown on monosaccharides namely arabinose, xylose, glucose, galactose and fructose than those on disaccharides or polysaccharides, Figure 1, (Rajoka and Malik, 1997). CBH was mainly present in the intracellular preparation. The organism synthesized CBH to a measurable level from disaccharides, otherwise only basal level of this enzyme was produced. Similarly FPase was produced in very small quantity below the detection limits of the assays or could not be quantified due to the presence of soluble sugars. Following growth on mono- and di-saccharides, cell extract containing cellobiohydrolase was collected and assayed for this activity. Carbon sources which supported rapid growth measured as qS (specific substrate utilization rate), µ (h-1) QEP (productivity of extracellular proteins) and QIP (productivity of intracellular proteins) were the worst repressors of enzyme synthesis. Among polymeric substrates, a-cellulose supported the maximum activity, followed by kallar grass. Overall cellulosic substrates induced high level of enzyme activities. Among monosaccharides, only fructose synthesized cellobiohydrolase and supported the work of Nochureet al. (1993). Among disaccharides, cellobiose was the best inducer, followed by maltose and lactose. There was greater enhancement in cellobiohydrolase productivity (2.4-fold enhancement) following growth on a-cellulose over that obtained from cellobiose (Table 1). Additionally level of CBH varied over a 1.57- to 11-fold range with the non-inducing carbon sources namely, xylose, arabinose, fructose and inducing carbon sources namely maltose, cellobiose, cellodextrin and cellulose (1.2- to 2.38- fold) and showed inverse relationship with values of qS, µ, QIP and QEP. FPase got immobilized on cellulosic substrates (10 ± 0.25%) and could be successfully eluted with Tween 80; all values presented in different figures and tables have been compensated to include substrate-bound FPase. Similarly some cells got adsorbed on the surface of insoluble substrates. These cells contained CBH activity and the values of CBH have also been compensated to contain cell-bound CBH activity in Table 1. Effect of nitrogen sources on cellulase production In C. flavigena, the effect of nitrogen sources was tested by replacing NaNO3 in the medium with other compounds, maintaining equi-molar amount of nitrogen at 0.164 g/litre. The cultures were grown for 72 hrs, harvested and processed for enzyme assays (Table 3). Inorganic nitrogen sources including NH4 Cl, (NH4)2 SO4, NH4H2PO4 and organic nitrogen sources namely corn steep liquor and urea were the poor nitrogen sources of FPase synthesis. NaNO3, KNO3 and NH4NO3 were the best sources since C. flavigena possessed strong nitrate reductase activity which was induced by NO3 ions to an optimal level and repressed by free NH4 ions in the growth medium. Spiridonov and Wilson (1998) found that NH4 compounds are the most favourable nitrogen sources for protein and cellulase synthesis. Nakamura and Kitamura (1988) observed that polypeptone supported the maximum production of FPase and CBH by C. uda CB 4.
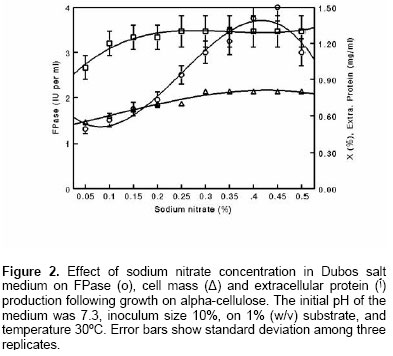
Effect of NaNO3 on exoglucanase synthesis The addition of NaNO3 increased cell mass and FPase synthesis when added to the basal medium (Figure 2). FPase was secreted at elevated levels (up to 2.5-fold higher) compared with control when alpha-cellulose was used as a cellulosic substrate. However, concentrations higher than 0.45% lowered the FPase activity to some extent but it was still higher than that present in the control. The yield was two- fold that obtained in mutant derivative of C. biazotea (Rajoka et al. 1998). The Qp levels of FPase on alpha-cellulose (87.5 IU/l/h) and kallar grass medium (79.5 IU/l/h) are greater than those reported on C. biazotea and its mutant derivative (18 IU/l/h and 21 IU/l/h; Rajoka et al. 1998) and other organisms (Spiridonov and Wilson, 1998; Kalogeris et al. 2003). During
growth of the organism on different cellulosic substrates, reducing sugars
accumulated (Table 4) in the growth medium as unmetabolized
principles. Those substrates which released more carbohydrates were stronger
repressors. The values of fermentation parameters with respect to substrate
utilization, namely maximum biomass and protein productivities, Qs and
Yx/s from a-cellulose, CMC and kallar
grass (Table 2). Indicated that maximum growth in terms
of dry cell biomass was significantly higher on a-cellulose
while minimum values was obtained following growth on CMC. The Yx/s of
Volumetric
productivity of CBH and growth parameters of the organism in Dubos-a-cellulose
medium when glucose was added (1, 2.5, 5 and 10 g/litre final concentration)
at the time of inoculation (Table 3), showed significant
decrease in the enzyme synthesis. All treatments had statistically significant
(p 0.05) influence on CBH productivity, substrate consumption
and cell# mass formation rates. It was found that glucose enhanced the cell
mass productivity, but suppressed CBH productivity and substrate consumption
rate. Mixed inductive or repressive effect has been observed in other organisms
(Ponce-Noyola and De Optimum
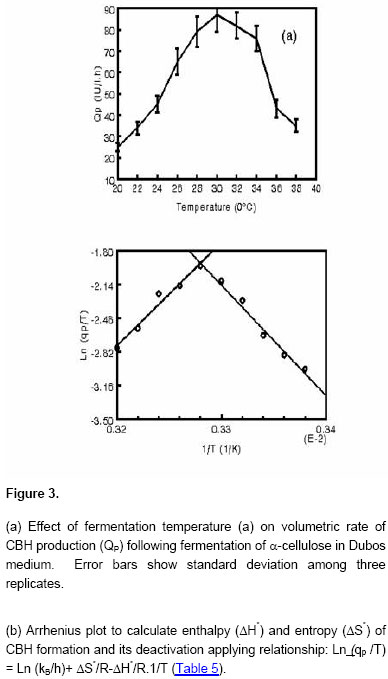
pH for production of CBH is 7.3 (results not shown). Temperatures higher
than Effect of temperature was shown by calculating activation enthalpy of CBH production graphically from Figure 3b by the application of Arrhenius approach (Aiba et al. 1973). The values of the activation enthalpy (Table 5) of CBH production (ΔH*= 86.9 k J/mol) are lower than that for glucose isomerase production reported by Converti and Del Borghi (1997). The phenomena responsible for thermal inactivation of enzyme is characterized by an activation enthalpy of ΔHD*= 92.6 k J/mol and is comparable with that for CBH production. This suggests that the productivities decline observed at high temperature could be due to the reversible denaturation of enzyme formed on Avicel medium. The activation entropy of CBH formation (0.095 k J/mol K) compares favourably with that of phytase formation (Converti and Dominguez, 2001). The activation entropy value of thermal inactivation (-0.498 kJ/mol. K) was also very low and had negative symbols which suggested that this inactivation phenomenon implied a little randomness/disorderness during the activated state formation and compared favourably with that of b-galactosidase formation (Rajoka et al. 2003). They further led to suggest that the phenomenon limiting CHB production could be enzymatic reaction/s under varying fermentation conditions as observed for other enzymes (Converti and Dominguez, 2001).
CONCLUDING REMARKS These studies led us to conclude that the carbohydrate and nitrogen sources play a vital role in the production of CBH or FPase by Cellulomonas. a-Cellulose was the best carbon followed by kallar grass but the former is an expensive substrate, therefore, kallar grass could be used for massl production of cellulases. In these studies, efforts were made to increase the enzyme production by manipulating above aspects only. The scope to increase the production by the use of genetical, biochemical and microbial engineering techniques to make use of full potential of this organism are to be studied. Gamma ray mutation followed by chemical mutagenesis have given mutant derivatives for improved cellulolysis (Gadgil et al. 1995; Rajoka et al. 1998) and will be used in further studies. Thermodynamic studies suggested that cells needed lower thermal energy for product formation and that the cell system exerted defence against thermal inactivation. REFERENCESNote: Electronic Journal of Biotechnology is not responsible if on-line references cited on manuscripts are not available any more after the date of publication. Copyright 2004 by Universidad Católica de Valparaíso -- Chile The following images related to this document are available:Photo images[ej04029f1.jpg] [ej04029f3a-b.jpg] [ej04029f2.jpg] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||

{kind=link}
{kind=link}
{kind=link}