 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Electronic Journal of Biotechnology, Vol. 7, No.3, December, 2004, pp. 295-301 RESEARCH ARTICLE Recombinant expression of marine shrimp lysozyme in Escherichia coli Enrique de-la-Re-Vega1,
Karina D. García-Orozco2, Sergio A. Calderón-Arredondo3,
María Gabriela Romo-Figueroa4, María
A. Islas-Osuna5, Gloria
M. Yepiz-Plascencia6, Rogerio R. Sotelo-Mundo*7 Code Number: ej04034 Shrimp Lysozyme (Lyz) is a key component of the antibacterial response as part of the innate defense in Crustacea; however, it has not been possible to purify this protein because of the very low amount present in the shrimp blood cells (hemocytes). In an effort to produce enough protein to study its function and biochemical properties we have overexpressed Lysozyme from marine shrimp (Penaeus vannamei) in E. coli. A bacterial protein expression system based on the T7 polymerase promoter was used. Although Lyz was produced as insoluble protein in inclusion bodies, its refolding led to an active protein with a yield of ~10%. Details of the protein recombinant expression techniques applied to this shrimp protein are presented. Keywords: crustacea, lysozyme, penaeideae, Penaeus vannamei, recombinant expression, refolding, shrimp. Abbreviations: ARTICLE Recombinant expression and refolding of proteins have provided a tool to study protein function, biochemical properties and structure where availability or yield precludes purification from the tissue of interest (Andersen and Krummen, 2002). Nowadays, many proteins are obtained by these techniques, in particular unidentified open reading frames from newly sequenced genomes, to identify the function of hypothetical polypeptides without apparent similarity to known proteins. Shrimp aquaculture is an important activity worldwide, providing employment and income to developing countries. This activity has been stricken by severe epizooties, both bacterial and viral. Therefore, there is a longstanding interest in understanding the mechanisms of antibacterial and recognition events that mediate the innate response of Crustaceans to microorganisms (Hultmark, 1996; Vargas-Albores and Yepiz-Plascencia, 2000; Yepiz-Plascencia et al. 2000). Efforts have focused on the purification and characterization of hemolymph (blood) proteins. Between them, Lysozyme is one of the earliest known antibacterial proteins, ubiquitous from phages to humans. In invertebrates, it is well known that the Lysozyme expression is regulated and responds to a bacterial challenge. Production of recombinant Lyz was possible after the cloning of the Lyz cDNA (Sotelo-Mundo et al. 2003). Furthermore, to this date it has not been reported the isolation or purification of shrimp Lyz from tissues or blood cells (hemocytes), which are the first line of innate defense in Crustacea. Bacterial expression is the most widely protein expression system used. However, it has important drawbacks that must be considered when expressing secreted or post-translationally modified proteins (De Bernardez Clark, 1998). Bacteria synthesize proteins in the cytosol where disulfide bonds and glycosylation do not occur. Therefore, the recombinant polypeptides that are not folded are stored as inclusion bodies. These apparent problems can be advantageous due to the high yields of denatured protein that may be synthetically refolded. Refolding involves formation of disulfides bonds and the exchange of scrambled incorrect disulfides by including small amounts of b-mercaptoethanol, or oxidized and reduced glutathione to increase the yield of properly folded proteins (Armstrong et al. 1999). One of the most common protein expression systems in Escherichia coli is based on the controlled expression of the gene of interest by the T7 RNA polymerase, under the control of the lac operon (Studier and Moffatt, 1986). In this system, the coding region of the protein to be expressed is inserted in a frame into the multiple cloning site provided in the expression vector. The coding region is amplified by PCR, and both insert and vector are digested with the corresponding restriction enzymes. If required, the plasmid vector is dephosphorylated to prevent recircularization during ligation to produce the recombinant vector (Sambrook et al. 2001). This paper describes details about the primer design, directional cloning, expression and activity of the shrimp Lyz protein. MATERIALS AND METHODSBacterial strains, plasmids and enzymes Escherichia
coli strain DH5a (Stratagene, Recombinant
techniques to make the shrimp Lyz expression construct were as described
(Sambrook et al. 2001) and based on the general cloning
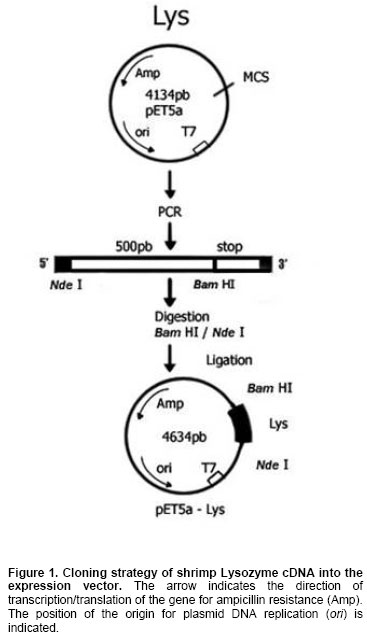
strategy shown in Figure 1.
It was prepared as follows: the Lysozyme coding region (Lyz DNA, GenBank
accession number AAL23948) was amplified by PCR with primers PV-Lyz-NdeI
5’GGAATTCCATATGTGCGACGCCAAGGTCTTC 3’ and PV-Lyz-BamHI
5’CGGGATCCTTAGAACGGGAAGACAGAG The template
for Lyz PCR was a shrimp hemocyte phagemid cDNA library (Sotelo-Mundo
et al. 2003). PCR was carried out in a 50 ml reaction containing The PCR
conditions were as follows: one cycle at The pET5a
vector and Lyz DNA were ligated overnight at Bacteria
were grown in LB broth containing ampicillin (100 mg/ml), kanamycin (15 mg/ml),
tetracycline (12.5 mg/ml) and chloramphenicol (34 mg/ml) for Lyz expression
in Rosetta Gami-LacZ. Culture in Fernbach flasks was done in an orbital
shaker at 250 rpm at The media
was centrifuged at 5000 x g for 5 min and the bacterial pellet was
washed in 0.9% NaCl (w/v) in a proportion of 20 ml of solution per g of pellet
and centrifugated at 5000 x g for 5 min. The bacterial cells were
resuspended with lysis buffer ( Refolding of Lyz from inclusion bodies Washed
inclusion bodies pellet were dissolved in an extraction buffer ( A search
for refolding conditions was done by testing the sixteen buffers reported
by Armstrong et al. (1999), by dilution of the inclusion
bodies solution, incubation at Buffer
#13 ( Shrimp
Lyz activity was determined by a modification of the turbidimetric method
(Shugar, 1952).In this approach, a suspension of Micrococcus
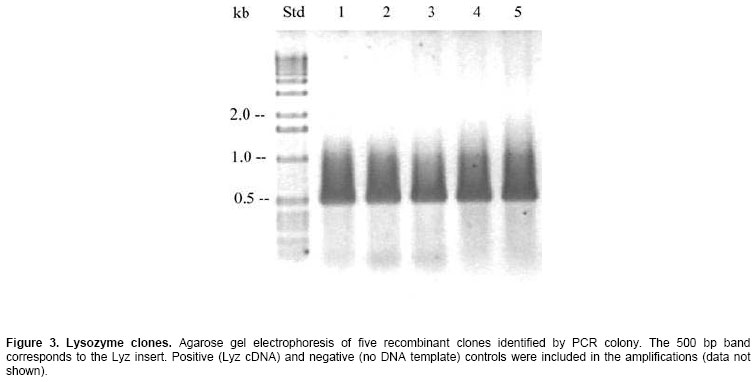
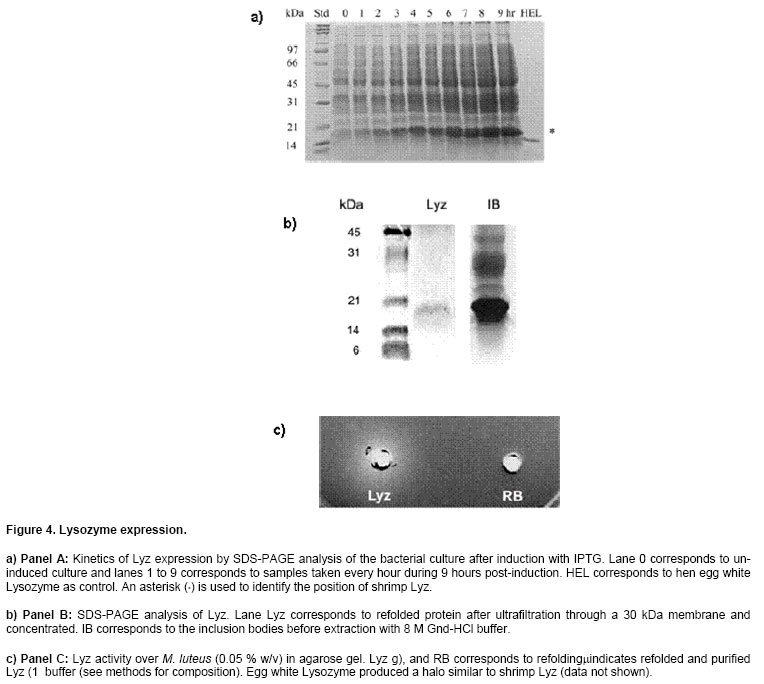
luteus (Sigma, St. Louis MO, Lyz cDNA cloning and its expression We obtained the recombinant construct pET5a-Lyz containing the coding region for the mature Lyz protein. Since the pET5a vector does not contain a blue/white selection system, we analyzed several putative clones by PCR colony and identified five of them containing the insert of ~500 bp that could correspond to Lyz (Figure 3). The identity of the 500 bp band was confirmed by DNA sequencing and corresponded to Lyz. Then, the construct pET5a-Lyz was re-transformed into the Rosetta Gami-lacZ strain for overexpression of the shrimp Lysozyme. Although the vector carries ampicillin resistance and only transformants should grow on a selective antibiotic media, transformed Rosetta Gami bacteria were also screened by PCR colony using primers PV-Lyz-BamHI (from Lysozyme) and T7 (from the vector) (data not shown) to identify the presence of pET5a-Lyz in such cells. Expression of Lyz was detected in the culture after 2 hrs of induction with IPTG by analysis of the bacterial pellet by SDS-PAGE, where the appearance of a ~17 kDa band indicates the synthesis of the recombinant Lyz (Figure 4, panel A). Soluble cytosolic and insoluble fractions of induced bacterial lysate showed that no Lyz activity was found in the soluble proteins, and by SDS-PAGE it was determined that Lyz expresses as inclusion bodies. From there, an inclusion body and refolding protocol was implemented. DTT was used to reduce any disulfide bonds during protein denaturation, however, it was diluted by dialysis on further steps and the protein was reoxidized by a combination of glutathione reduced/oxidized during refolding. Protein
concentration was monitored during the inclusion body isolation, refolding
and purification of Lyz, and a total yield of 10.04% was obtained, for a
total of 6.75 mg Lyz/lt of media. In Figure 4, panel B,
it can be observed that a band of ~17 kDa is the main component of the inclusion
bodies (IB lane). Refolding of disulfide-containing proteins is not trivial;
although it has been reported a factorial assay of sixteen conditions, some
of which help to refold denatured egg white Lysozyme (Armstrong
et al. 1999). In brief, the sixteen buffers were tested on Lyz inclusion
bodies, and refolding was identified by enzymatic activity on the solution.
We tested such conditions and found that one of them produced active refolded
Lyz with better yield (buffer #13: Bacterial expression is challenging, although it has the advantage of a low cost compared to higher eukaryotic systems such as baculovirus or mammalian cell culture (De Bernardez Clark, 1998). In the event of expressing translationally-modified proteins, once the refolding protocol has been established, it is suitable to be scaled up since it involves operations such as dialysis and may be substituted by diafiltration with membranes. Factors such as media composition or scale-up to a fermentor may be more critical to increase expression yield. Other vector seems not to be a critical factor in Lyz expression, although other vectors such as pET11a which provides a tighter expression control may increase the yield. Other Lyz refolding is not trivial, being refolding a critical step. Disulfide bond formation seems to be the most challenging step, and buffers that contain combination of thiol oxidant/reducer such as oxidized/reduced glutathione or cystine/cysteine are valuable additives during refolding (Armstrong et al. 1999). Manduca sexta Lysozyme was refolded also from E. coli inclusion bodies (Lopez-Zavala et al. 2004). In conclusion, we have been able to obtain recombinant Lyz in sufficient amounts for further biochemical and structural studies. In our experience, recombinant expression should be used in addition to chromatographic purification of proteins, having the cDNA available or by making use of sequences available in Genbank. Other advantage of the recombinant expression is that site-directed mutagenesis allows investigating the function of specific residues involved in catalysis or as in Lyz, antibacterial activity. It can be anticipated that multiple biotechnological applications will be implemented upon application of recombinant expression of proteins from organisms such as the marine crustacean. ACKNOWLEDGMENTSWe thank
Sandra de Note: Electronic Journal of Biotechnology is not responsible if on-line references cited on manuscripts are not available any more after the date of publication. Copyright 2004 by Universidad Católica de Valparaíso -- Chile The following images related to this document are available:Photo images[ej04034f1.jpg] [ej04034f3.jpg] [ej04034f2.jpg] [ej04034f4a-c.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}