 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
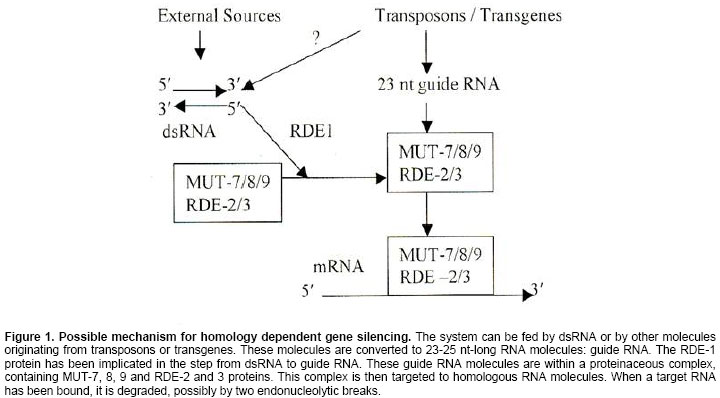
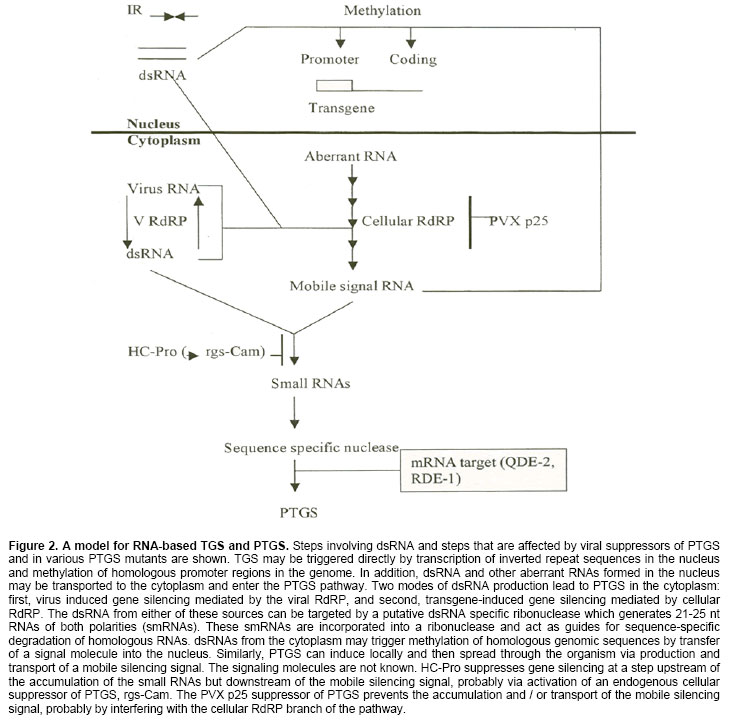
Electronic Journal of Biotechnology, Vol. 7, No.3, December, 2004, pp. 321-332 REVIEW ARTICLE Mechanisms and roles of the RNA-based gene silencing Snehasis Jana1, Chiranjib Chakraborty*2, Shyamsundar Nandi3 1Department
of Biochemical Engineering and Biotechnology
Indian Institute
of Technology
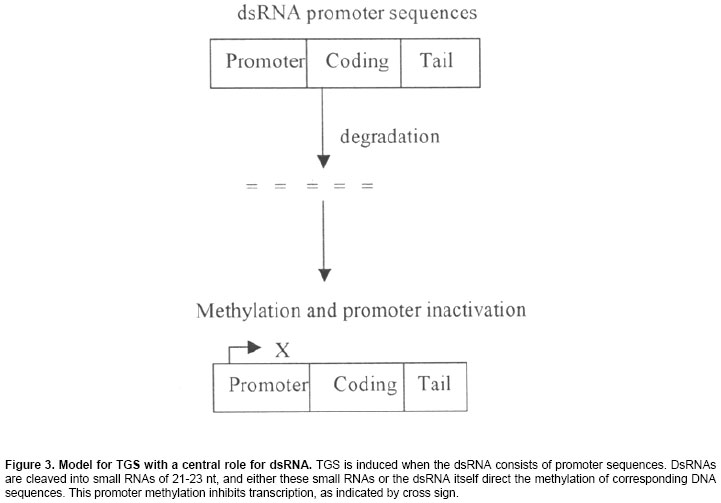
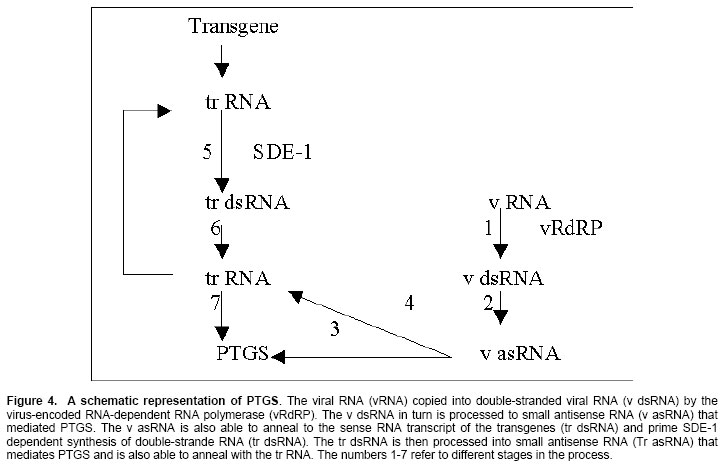
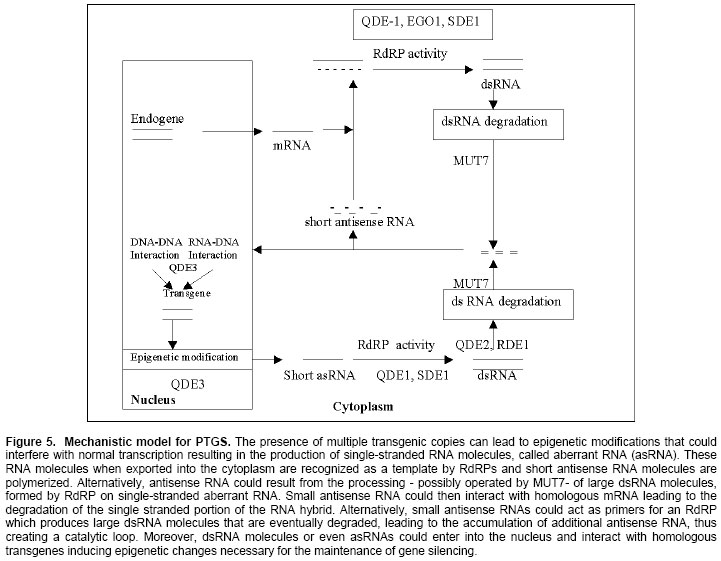
New Delhi, India ABSTRACT RNA silencing is a remarkable type of gene regulation. This process has been found to occur in many different organisms such as plants (co-suppression), fungi (quelling), and animals (RNA interference; RNAi). Double-stranded RNA (dsRNA) is a potent trigger in RNA silencing mechanisms operating in a wide range of organisms. This mechanism recognizes dsRNA and processes them into small 21-25nt RNAs (smRNAs). Small RNAs can guide post-transcriptional degradation of complementary messenger RNAs and in plants, transcriptional gene silencing is occurred by methylation of homologous DNA sequences. In plants, it serves as an antiviral defense, and many plant viruses encode suppressors of silencing such as helper component-proteinase of potyviruses (HC-Pro) and the p25 protein encoded by potato virus X (PVX). HC-Pro acts by preventing accumulation of smRNAs that provide specificity determinant for homologous RNA degradation, but p25 viral protein acts by targeting the mobile silencing signal. The encouraging view is that RNA silencing is part of a sophisticated network of interconnected pathways for cellular defense and development and that it may become a powerful tool to manipulate gene expression experimentally. Keywords: double stranded RNA, post-transcriptional gene silencing, RNA interference, transcriptional gene silencing. ARTICLERNA silencing was first discovered in transgenic plants where it was termed co-suppression or post-transcriptional gene silencing (PTGS). There is a sophisticated machinery for homology-dependent gene silencing (HDGS) (Figure 1) that seems essentially common between algae, fungi, plants and animals (Baulcombe, 1996; Alvarado and Newmark, 1999). At least four entirely independent lines of research led to this realization: transgene-dependent gene silencing in plants (co-suppression), quelling in fungi, RNA interference (RNAi) in diverse animals (Montgomery and Fire, 1998; Vaucheret et al. 1998;), and the silencing of transposable elements (Cogoni and Macino, 1999). A key feature uniting the RNA silencing pathways in different organisms is the importance of double-stranded RNA (dsRNA) as a trigger or intermediate. The dsRNA is cleaved into the small interfering RNA (siRNAs) that is ~ 21-25 nucleotides and these can trigger the degradation of homologous RNAs in the cytoplasm (PTGS) and de novo methylation of homologous DNA in the nucleus (Wassenegger et al. 1994; Carthew, 2001). At the genome level, RNA can induce the epigenetic modification of homologous DNA sequences through the process of RNA-directed DNA methylation (RdDM), that has only been demonstrated in plants. Two distinct gene silencing phenomena are observed in plants: transcriptional gene silencing (TGS), which involves decreased RNA synthesis because of promoter methylation (Cogoni and Macino, 2000; Wassenegger et al. 1994), and post transcriptional gene silencing (PTGS), which involves sequence specific RNA degradation (Fire, 1999). PTGS is induced by deliberate or fortuitous production of dsRNA (Couzin, 2002). RNA interference (RNAi) is also a gene-silencing phenomenon triggered by dsRNA and involves the generation of smRNAs (Hamilton and Baulcombe, 1999; Matzke et al. 2001). An important aspect of RNA silencing in plants is that it can be triggered locally and then spread via a mobile silencing signal (Vance and Vaucheret, 2001). The signaling molecule is not known but is expected to contain a nucleic acid component to account for the sequence specificity. Systemic spread of silencing also occurs in the organisms; through the mechanism may not be the same as in the plants (Ngo et al. 1998; Plasterk and Ketting, 2000). In this review we describe mechanisms of two distinct gene-silencing phenomena and consider their roles in host defense system and in development. A model of RNA silencingRNA-directed DNA methylation (RdDM) and post-transcriptional gene silencing/RNA interference (PTGS/RNAi) are both triggered by dsRNAs that are cleaved by RNAseIII-type enzymes (i.e. Dicer, Caf) into small interfering RNAs (siRNAs), probably in both the nucleus and the cytoplasm (Voinnet, 2002; Zamore, 2002). In the cytoplasm, the siRNAs serve as guides for endonucleolytic cleavage of homologous mRNA in association with the RNA-induced silencing complex (RISC). DsRNA can trigger RdDM. RNA triggering of RdDM might either interact with the chromodomain of chromomethylase (CMT) and guide it to the homologous DNA sequence, that might attract a de novo DNA methyltransferase (DNMT). DsRNAs can be made by transcribing through inverted DNA repeats (IR) or by the activity of cellular RNA-dependent RNA polymerase (cRdRP) acting on 'aberrant' (prematurely terminated and / or lacking polyadenylation) RNA templates synthesized from single copy (SC) genes in the nucleus or generated in the cytoplasm by RISC cleavage of mRNA (Hamilton et al. 2002; Llave et al. 2002). Replicating RNA viruses produce dsRNA by means of a viral RdRP (vRdRP). Recent studies of two plant viral suppressors (Hamilton and Baulcombe, 1999), the helper component-proteinase (HC-Pro) of potyviruses and the p25 protein encoded by potato virus X (PVX), represent two different viral strategies to suppress silencing (Figure 2). Hc-Pro is a highly effective suppressor of silencing that can enhance the accumulation of a broad range of unrelated plant viruses (Vance and Vaucheret, 2001). It prevents both virus-induced gene silencing (VIGS) and transgene-induced RNA silencing, and it reverses an already established RNA silencing of a transgene, but fails to eliminate the mobile silencing signal (Mallory et al. 2001) as assayed by grafting experiments. PVX p25 is much less effective in blocking silencing than HC-Pro, and it appears to target and interfere with systemic silencing (Voinnet et al. 2000). The exact nature of the systemic silencing signal is unknown, but it involves either dsRNA or a special class of siRNAs (Tanget al. 2003; Timmons and Fire, 1998). Transcriptional gene silencingRNA-dependent DNA methylation (RdDM) was first discovered with viroids (Angell and Baulcombe, 1997; Jones et al. 2001), which are plant pathogens consisting of non-coding, highly base paired, rod shaped RNA several hundred nucleotides in length (Beclin et al. 2002; Fagard et al. 2000). RdDM results in dense methylation at most symmetrical and non-symmetrical cytosines within the region of homology between the inducing RNA and the target DNA. DNA targets as short as 30bp can be modified. RdDM provides an alternate means to induce the sequence specific methylation observed in both PTGS and TGS. The ability of viroids and RNA viruses, which produce dsRNA replication intermediates, to trigger RdDM suggested a general requirement for dsRNA in this process (Cox et al. 1998). A dsRNA transcribed from an IR containing promoter sequences is able to trigger de novo methylation and silencing of homologous promoter in trans. The promoter dsRNA is degraded to smRNAs, indicating that it is entered the same degradation pathway as dsRNAs involved in PTGS. Thus, dsRNA provides a common molecular link between RdDM, which can lead to TGS if promoter sequences are involved, and the RNA degradation step of PTGS (Kasschau et al. 2003; Llave et al. 2002). The mechanism of RdDM is unknown but is assumed to involved RNA-DNA interactions based on sequence homology. The minimal DNA target size for RdDM of 30 bp opens the possibility that the 21-25 nucleotides (nt) RNA degradation products of dsRNA could be responsible for directing de novo methylation (Mallory et al. 2002; Montgomery and Fire, 1998). The smRNAs could conceivably guide the DNA methyltransferase to unmodified homologous DNA sequences. The methylation of promoter sequences (Mette et al. 2000) usually results in promoter inactivation (Figure 3), probably by histone deacetylation and chromatin condensation (Wassenegger et al. 1994). RNAinterference is a conserved process in which double-stranded RNA is processed into 21-25 nucleotide siRNAs that trigger posttranscriptional gene silencing. In addition, plants display a phenomenon termed RNA-directed DNA methylation (RdDM) in which DNA with sequence identity to silenced RNA is de novo methylated at its cytosine residues (Cao et al. 2003). This methylation is not only at canonical CpG sites but also at cytosines in CpNpG and asymmetric sequence contexts. Cytosine DNA methylation silences harmful DNAs such as transposons and retroviruses. RDR2, DCL3, and AGO4 are homologous to proteins involved in RNA silencing, a conserved genome defense mechanism that uses short, 21- to 25-nucleotide silencing-induced RNAs (siRNAs) to direct posttranscriptional messenger RNA (mRNA) destruction and also to cause chromatin-level gene silencing (Simon et al. 2004). siRNAs are processed from double-stranded RNA by Dicer, an RNaseIII-RNA helicase, and their synthesis often requires an RNA-dependent RNA polymerase. RNA thus appears to be a general means of targeting de novo DNA methylation, which may indicate how sequence-specific gene silencing is established in a variety of epigenetic phenomena. Several studies suggest the possible involvement of DNA triplet helix structure in the process of RdDM. These unusual structures might attract DNMT (Tang et al. 2003; Vance and Vaucheret, 2001). Another possible candidate methyltransferase is the so-called chromodomain containing methyltransferases i.e. chromomethylase, that has been found only in plants. Chromodomains are believed to mediate interactions between chromatin regulatory proteins. Small RNAs might interact with a chromomethylase through the chromodomain to direct methylation of homologous DNA sequences (Sijen et al. 2001; Matzke, 1995). Post-transcriptional gene silencing (PTGS)Post-transcriptional gene silencing (Sijen et al. 2001; Cogoni and Macino, 2000) in plants, quelling in fungi, and RNA interference in animals: are responses to various types of foreign nucleic acid including viruses, transposons, transgenes and dsRNA (Dalmay et al. 2000). These processes represent natural defence mechanisms against viruses and transposons (Ketting and Plasterk, 2000). The silencing effect can be transmitted systemically from a silenced regions (stock) to unsilenced region (scions) in plants and so that the silencing signal can travel over a long distance through the plant's vascular system. A remarkable characteristic of PTGS in plants and fungi is that diffusible trans-acting molecules mediate it, which is able to mediate gene silencing between nuclei in heterokaryotic strains. The small sense and antisense RNAs associated with silencing derive from cleavage of dsRNA and both polarities are thought to incorporate into a ribonuclease complex, serving as a guide to find homologous target RNAs. Small RNAs accumulate during both virus and transgene-induced gene silencing, indicating convergence of these two branches of silencing before the formation of the sequence-specific ribonuclease (Fagard et al. 2000). In plants, dsRNA that triggers PTGS can be produced in the nucleus by transcription through inverted repeat (IRs) or through the action of RdRP, which is postulated to use pre-existing dsRNA or 'aberrant'sense RNAs (Montgomery and Fire, 1998; Li et al. 2000) as templates for the synthesis of antisense RNAs (Figure 4). Several reports which suggested that PTGS can be induced by dsRNA, that small RNAs are produced and that cognate DNAs become methylated suggested that TGS and PTGS are mechanistically linked (Mallory et al. 2002; Matzke et al. 2001). The cellular machinery does not seem to discriminate between dsRNAs consisting of coding and promoter sequences, and both are cleaved into small RNAs. When homologous mRNAs are present, the small antisense RNAs may anneal and guide degradation of these mRNAs but prior to degradation, an RNA-dependent RNA polymerase (RdRP) may elongate some of the guide RNAs on the mRNA, after which the dsRNA part is degraded (Hunter, 1999; Llave et al. 2000; Kasschau et al. 2003). Mechanism of PTGSThe cloning of genes coding for cellular components of the PTGS (Timmons and Fire, 1998) machinery in different system such as Neurospora crassa, Caenorhabditis elegans and Arabidopsis thaliana has demonstrated the existence of a common genetic basis PTGS. RdRP plays a key role in PTGS (Smardon et al. 2000; Tabara et al. 1999). Initial models proposed that copy antisense RNA (cRNA) could be produced by an RdRP on a sense transgenic mRNA, usually referred to as aberrant RNA (aRNA) and that is able to mediate gene silencing in plants and fungi through diffusible signal (Vaucheret et al. 1998). It has been suggested that such aRNA, produced as a consequence of an epigenetic alteration of transgenic loci (Plasterk and Ketting, 2000), could be incorrectly terminated covalently modified and / or complexed with specific proteins (Table 1).
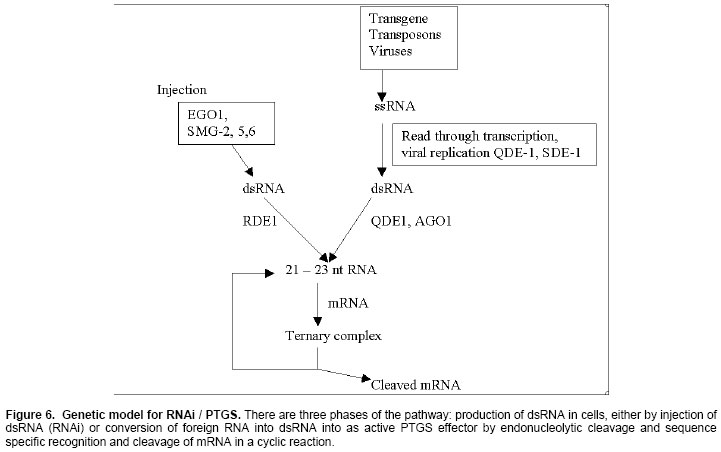
The RdRPs are required even RNAi has led to the hypothesis that RdRP template in transgenic-induced PTGS could be dsRNA produced directly by inverted repeat transgenic loci. RdRP in fungi and plants is necessary to produce large dsRNA molecules on single stranded transgenic aberrant RNAs (Dalmay et al. 2000). However, small antisense RNAs could act as primers to direct RdRPs to single stranded homologous mRNA with the large dsRNA molecules formed, thereby being immediately processed into short antisense/sense RNAs (Brantl, 2002; Grishok et al. 2001) that initiate a new cycle of antisense RNA annealing, RdRP-mediated dsRNA synthesis and dsRNA degradation (Figure 5). A model for RNA interference and PTGSFigure 5 represents a speculative model that draws upon the genetic evidence. Promiscuous transcription of transgenes or transposons produces RNA that serves as a template for RdRP (Bosher et al. 1999). RdRP synthesizes complementary RNA that consequently base paires with its template. Alternatively, dsRNA is produced, during viral infection either as a replicative intermediate or as a product of overlapping transcription (Angell and Baulcombe, 1997; Carmell et al. 2002). Once produced, dsRNA might replicate itself using RdRP in association with dephosphorylation SMG-2 (Mourrain et al. 2000), allowing an initial amount of dsRNA to increase its abundance and resist being diluted by cell growth (Doench et al. 2003). In parallel, some of the dsRNA is uniformly cleaved at specific sites by a dsRNA endonuclease to form a non-overlapping set of 21-23 nt RNA fragments (Hamilton et al. 2002). A newly cleaved RNA fragment remains associated with the nuclease to form a stable ribonucleo protein (RNP) complex (Figure 6). The RDE-1, QDE-2 and AGO1 factors probably facilitate formation of the RNP complex (Fire et al. 1998; Guo and Kempues, 1995; Elbashir, et al. 2001). An mRNA molecule would specifically associate with the RNP complex on the basis of its sequence complementarity with the antisense strand of the guide RNA (Ketting et al. 1999; Martinez et al. 2002; Miyagishi et al. 2003). This would require displacement of the sense strand. The mRNA might then be cleaved at two sites to generate 21-23 base pair dsRNA within the RNP complex (Reinhart et al. 2000; Seggerson et al. 2002). If a helicase activity unwinds the dsRNA isoform, the RNP complex would be free to carry out another round of the base paring and cleavage. Thus the reaction is cyclic (Mallory et al. 2002; Morel et al. 2002; Mourelatos et al. 2002). Roles of RNA silencingRNA based silencing mechanisms which are effective at the genome level and in the cytoplasm, are able to combat parasitic sequences that have an RNA genome (RNA viruses) or a dsRNA replication intermediate (Zamore, 2001). The host defence function of RNA-mediated silencing is demonstrated by the increased sensitivity of Arabidopsis PTGS mutants to some viruses and the mobilization of transposons in RNAi mutants (Reinhart et al. 2000; Wargelius and Ellingsen, 1999) of C. elegans and in the Mut-6 mutant of Chlamydomonas. The native function of PTGS is most likely in providing resistance to virus infection. It has been found that many viruses are potent inducers of PTGS and viruses encode factors that inhibit this response of the plant (Voinnet, 2002; Walters and Jelinek, 2002). RNA silencing is an evolutionarily conserved surveillance systemthat occurs in a broad range of eukaryotic organisms. In plants,RNA silencing acts as an antiviral system; thus, successfulvirus infection requires suppression of gene silencing (Shuey et al. 2002; Zilberman et al. 2003). A numberof viral suppressors have been identified so far. We describe one of viral suppressor protein p19 of Cymbidium ringspot virus (CymRSV), which inhibitsRNA silencing via its small RNA-binding activity in vivo (Lakatos et al. 2004). SmallRNAs bound by p19 in planta are bona fide double-stranded siRNAsand they are silencing competent in the in vitro (Silhavy et al. 2002) RNA-silencingsystem. p19 also suppresses RNA silencing in the heterologousDrosophila in vitro system (Grishok et al. 2001) by preventing siRNA incorporationinto RISC. During CymRSV infection, p19 markedly diminishesthe amount of free siRNA in cells by forming p19-siRNAcomplexes, thus making siRNAs inaccessible for effector complexesof RNA-silencing machinery. Furthermore, reported resultsalso suggest that the p19-mediated sequestration of siRNAs invirus-infected cells blocks the spread of the mobile, systemicsignal of RNA silencing (Vance and Vaucheret, 2001). MicroRNAs (miRNAs) are single-stranded 21-25 nucleotide RNAs that regulate gene expression. miRNAs are cut from longer stem-loop precursors by Dicer and other members of the ribonuclease III family (Grishok et al. 2001; Ketting et al. 2001). Unlike siRNAs, the precursors to miRNAs are usually transcribed from non-coding genes separate from the genes they regulate (Hutvagner et al. 2001). In plants, most miRNAs are single-stranded mature miRNAs and the complementary strands are usually degraded. Perhaps p19 disrupts normal plant development because it traps miRNAs in a double-stranded state, preventing them from maturing into the single stranded form required for their function as developmental regulators (Papp et al. 2003). In plants, PTGS has been strictly linked to RNA virus resistance mechanisms. Plant RNA viruses are in fact both triggers and targets for PTGS, and display increased sensitivity to viral infections (Silva et al. 2002). The finding that mutations in the C. elegans mut-7 gene affect both RNAi and transposon silencing in the germline indicates clearly that RNAi is a mechanism implicated in transposons control (Zamore, 2002). Silva et al (2002) reported recently that nodaviral infection triggers an RNAsilencing-based antiviral response (RSAR) in Drosophila, whichis capable of a rapid virus clearance in the absence of expressionof a virus-encoded suppressor. Theyreported that vaccinia virus and human influenza A, B, and C viruseseach encode an essential protein that suppresses RSAR in Drosophila. Their findingsprovide evidence that mammalian virus proteins can inhibit RNAsilencing, implicating this mechanism as a nucleic acid-basedantiviral immunity in mammalian cells. Many mutants impaired in RNA silencing in plants, fungi, and animals have no obvious phenotype, suggesting that the corresponding genes, as well as silencing itself, are dispensable for normal development (Shuey et al. 2002). In contrast, other silencing mutants exhibit developmental abnormalities and are infertile, such as Ago1, caf/sin1/sus1 in arabidopsis, ego-1, mut-7 in C. elegans, suggesting that these genes play independent roles in development and silencing (Miyagishi et al. 2003). Furthermore, plants that express high levels of the viral silencing suppressor HC-Pro or that over express that endogenous suppressor rgs-Cam also show abnormal development. This raises the possibility that the suppression of RNA silencing mediated by these proteins works via interactions with factors that have dual role in silencing and development (Llave et al. 2002; Finnegan et al. 2003). Thus, development appears to be another of the interconnected network of pathways of which RNA silencing is a part. With our present understanding of the process, however, it is clear that RNA silencing has enormous potential for engineering control of gene expression, as well as a tool in functional genomics. CONCLUDING REMARKSThe involvement of dsRNA and DNA methylation in both TGS and PTGS type of silencing is increasingly recognized. The generation of RdDM mutants in Arabidopsis and the identification of endogenous DNA-target sequences will help to establish the actual mechanism and natural roles of this process. RNA helicases are potential candidate for the RdDM machinery as it is likely that RNA-DNA paring requires a ssRNA that is complementary to target DNA. We do not know the details of the silencing mechanism, especially not of the way transposons feed into it. It seems that copy number is bad news. We are proposing that the dsRNA may be a red signal for multicopy DNA. As soon as multiple copies are found in the genome, there is bound to be transcription of one strand here and of another strand there, allowing formation of dsRNA. This field is still in its romantic phase. Several questions will remain. Is the silencing effect replicated, and how? How does the guide RNA can find its target and ear mark it for degradation? How does the trigger RNA migrate in plants and animals? How does the mechanism feed back into the nucleus to silence gene transcription? How does an epigenetic modification influence the initiation and maintenance of PTGS and RdDM mediated TGS? Thus, genetics as well as biochemistry will need to be more exercised to resolve all these questions. ACKNOWLEDGEMENTSWe thank Ch. Ushashri, Nidhi Gupta and Rupali Walia for helpful comments on the manuscript.
Note: Electronic Journal of Biotechnology is not responsible if on-line references cited on manuscripts are not available any more after the date of publication. Copyright 2004 by Universidad Católica de Valparaíso -- Chile The following images related to this document are available:Photo images[ej04037f4.jpg] [ej04037f5.jpg] [ej04037f1.jpg] [ej04037f6.jpg] [ej04037f3.jpg] [ej04037f2.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}