
|
Electronic Journal of Biotechnology
Universidad Católica de Valparaíso
ISSN: 0717-3458
Vol. 8, Num. 1, 2005, pp. 35-42
|
Electronic Journal of Biotechnology, Vol. 8, No.1, April, 2005
RESEARCH ARTICLE
Production of an extracellular keratinase from Chryseobacterium sp.
growing on raw feathers
Adriano Brandelli*1, Alessandro Riffel2
1Laboratorio de Bioquímica e Microbiologia Aplicada,
Departamento de Ciência de Alimentos, ICTA,
Universidade Federal do Rio Grande do
Sul,
91501-970 Porto Alegre, Brasil,
Tel: 551 3316 6249,
Fax: 551 3316 7048,
E-mail: abrand@ufrgs.br
2Laboratorio de Bioquímica e Microbiologia Aplicada,
Departamento de Ciência de Alimentos, ICTA,
Universidade Federal do Rio Grande do
Sul,
91501-970 Porto Alegre, Brasil,
Tel: 551 3316 6249,
Fax: 551 3316 7048,
E-mail: ariffel@esalq.usp.br
*Corresponding author
Financial support: This work was supported by CNPq and FAPERGS.
Received September 28, 2004 /
Accepted January
13, 2005
Code Number: ej05006
ABSTRACT
The strain Chryseobacterium sp. kr6 shown to be useful for biotechnological
purposes such as hydrolysis of poultry feathers and de-hairing of bovine
pelts. The effect of temperature, initial pH and media composition on protease
production by this keratinolytic strain was studied. The enzyme was produced
between 25 and 37ºC, with maximum activity and
yield at 30ºC.
When protease production was tested in media with different initial pH, maximum
activity was observed when cultivation was carried out at 30ºC and initial pH ranging from
6.0 to 8.0. Higher activity was observed when feathers or feather meal were
used as growth substrates, followed by soybean meal. The addition of carbohydrates
or surfactants to feather broth resulted in decrease in keratinolytic activity.
Keywords: bacteria, enzyme, keratin, protease.
Proteolytic enzymes are largely used in the industry for biotechnological
applications involving the hydrolysis of protein substrates. Proteases constitute
an important fraction of the global enzyme sales, and a relevant part of this
market is accounted by bacterial proteases (Rao
et al. 1998).
Bacterial keratinases are of particular interest because of their action on
insoluble keratin substrates, and generally on a broad range of protein substrates
(Lin et al. 1995). These enzymes have been studied for de-hairing
processes in the leather industry (Raju et al. 1996) and
hydrolysis of feather keratin (Lin et al. 1995), which is
a by-product generated in huge amounts by the poultry industry. Discarded feathers
are currently used to produce feather meal through thermal processing, resulting
in a low nutritional value product (Wang and Parsons, 1997).
Feather hydrolysates produced by bacterial keratinases have been used as additives
for animal feed (Williams et al. 1991). In addition, keratin
hydrolysates have potential use as organic fertilizers, production of edible
films and rare amino acids (Dalev and Neitchev, 1991; Choi
and Nelson, 1996).
Cultivation conditions are essential in successful production
of an enzyme, and optimization of parameters such as pH, temperature and media
composition is important in developing the cultivation process. Despite all
the work that has been done on production of proteolytic enzymes, relatively
little information is available on keratinases (Wang and Shih,
1999). This is even more the case for keratinases of Gram-negative bacteria
(Sangali and Brandelli, 2000). The Chryseobacterium sp.
strain kr6 was isolated from waste of a poultry industry and was capable to
completely degrade chicken feathers. Its extracellular keratinase is a metalloprotease
with great potential for biotechnological applications (Riffel
et al. 2003a). This work describes the effect of temperature, initial pH
and substrates on keratinase production by Chryseobacterium sp. kr6
during growth on native feathers.
MATERIALS AND METHODS
Bacterial strain and
media
The bacterium Chryseobacterium sp. strain kr6 (LBM
9006) was isolated and characterized as described by Riffel et
al. (2003a). Brain heart infusion broth (BHI, Difco) medium was used for
maintenance of strain with 20% (v/v) glycerol at -20ºC.
Enzyme activity
and protein determination
The proteolytic activity was monitored as describe previously
(Sangali and Brandelli, 2000). Briefly, 100 μl of enzyme
extract was added to 400 μl of 10 mg ml-1 azokeratin in 0.1
M phosphate pH 8.0. The reaction mixture was incubated
for 15 min at 50ºC and
then stopped by addition of trichloroacetic acid to reach a final concentration
of 10% (w/v). Absorbance at 440 nm was measured after centrifugation at 10,000 g for 5 min. One unit of activity
was the amount of enzyme that caused a change of absorbance of 0.01 at 440
nm in 15 min at 50ºC.
Azokeratin was synthesized as described by Riffel et al. (2003b).
The protein concentration was measured by the Folin phenol reagent method (Lowry
et al. 1951).
Growth conditions
The inoculum was prepared by streaking a feather meal agar
(10 g l-1 feather meal, 15
g l-1 agar, 5
g l-1 NaCl, 0.4
g l-1 Na2HPO4, 0.3 g l-1 NaH2PO4)
plate with cells from a stock culture of kr6 strain. A single colony was transferred
to 10 ml of feather meal broth (FMB; 10 g l-1 feather meal, 5
g l-1 NaCl, 0.4
g l-1 Na2HPO4, 0.3 g l-1 NaH2PO4),
incubated at 30ºC until
the cell density reached about 108 cells ml-1. Then 1.0
ml was transferred to 500 ml Erlenmeyer flasks containing 100 ml of feather
keratin broth (FKB; washed, whole feathers are used instead of feather meal)
and cultivated in a shaker at 180 cycles min-1 for desired times.
Bacterial growth was monitored by viable cell counts (CFU ml-1)
and related to biomass dry weight with a calibration curve. The culture supernatants
were used for assays of proteolytic activity.
The influence of temperature on growth and production of protease
was studied at 25ºC, 30ºC, 37ºC and 42ºC.
Protease production was also investigated in FKB medium with initial pH adjusted
to 5.0, 6.0, 7.0, 8.0 and 9.0. The cultivation was carried out at 30ºC.
When different protein sources were used as substrates, a
concentration of 10 g l-1 was
used, replacing the raw feathers. The effect of carbohydrates on keratinase
production was tested by adding 10 g l-1 of
glucose, sucrose or lactose to FKB. The effect of surfactants was tested by
the addition of 10 g l-1 of Tween 80 or Triton
X-100.
All experiments were done in triplicate.
Kinetic parameters
Kinetic data were analyzed as described elsewhere (Sinclair
and Cantero, 1990; Doran, 1995). The maximal specific
growth rate (μmax) was calculated from the equation:
μ = dX/Xdt
where X is the cell concentration in g l-1 and
t the time (h).
Specific production rates (qP), defined
as the amount of activity produced per gram of cell dry mass and per hour (U
g-1 h-1) were calculated during growth from the relation:
qp = dP/Xdt
where P is the activity expressed in U l-1, X the
biomass concentration on g l-1 and t the time (h).
RESULTS
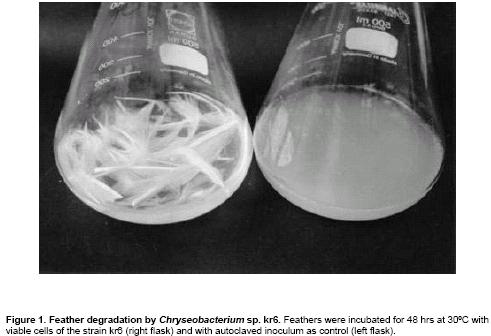
The strain of Chryseobacterium sp.
kr6 grew well and completely degraded chicken feathers in the medium (Figure
1). This intense feather-degrading activity was achieved in the range of
25-37ºC and
with initial pH adjusted from 6.0 to 8.0. Similar growth curves were observed
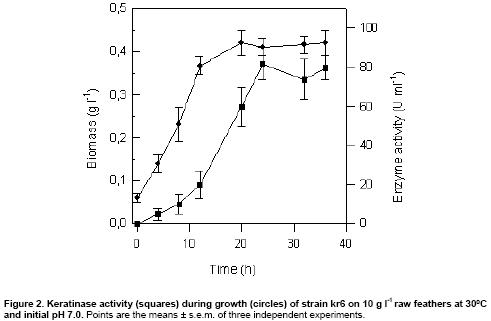
within this range of temperature and pH. A typical curve is shown in Figure
2. The stationary growth phase was reached after 24 h, coinciding with
maximum proteolytic activity. In all cases, the pH values increased up to 8.3
during cultivation, and maximum specific production rate was achieved at 24
h, coinciding with the end of exponential growth phase. No important growth
neither proteolytic activity were observed during cultivation of the strain
at 42ºC or higher and pH 9.0 or higher
(not shown).
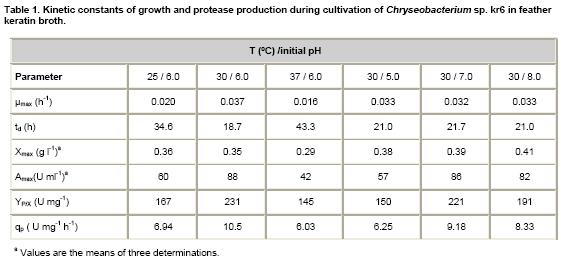
The maximum specific growth rate (μmax), the
doubling time (td), the yield of specific product formation (YP/X),
and specific production rate (qp), calculated at the final
growth phase are shown in Table
1. Maximum enzyme activity
and enzyme yields were observed at 30ºC and
pH ranging from 6.0 to 7.0. Maximum biomass was obtained with more alkaline
pH values (Table 1). Although the production pattern
of keratinase was similar for the different pH values tested, lower activities
were observed at pH 5.0. The qp values were higher at 30ºC and pH 6.0-7.0.
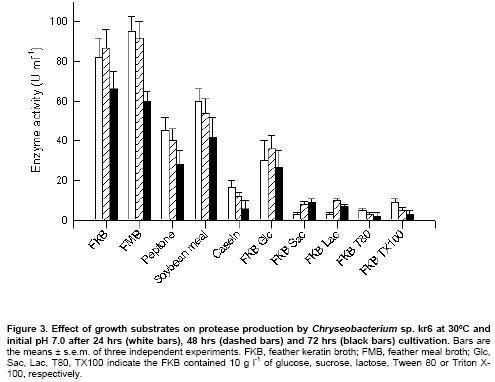
The effect of various substrates on keratinase production
was investigated. Production of keratinase activity was similar when the strain
kr6 was grown in raw feathers or feather meal, but decreased with other proteinaceous
substrates (Figure 3). The FKB medium was supplemented
with carbohydrates and the production of keratinase was then evaluated. The
addition of glucose, and markedly sucrose or lactose, resulted in strong inhibition
of keratinase synthesis (Figure 3). Other additives such
as the surfactants Tween 80 and Triton X-100 also caused reduction in protease
yields of about 90% and 75%, respectively.
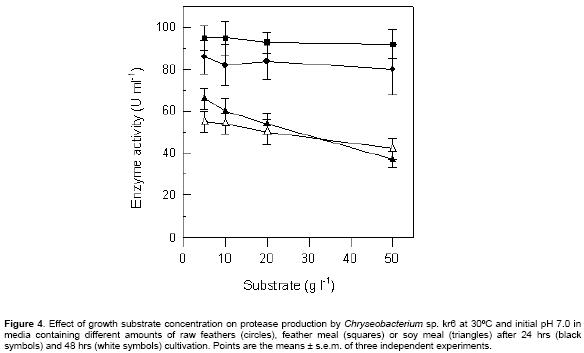
The effect of concentration of three growth substrates on
keratinase production was tested. Keratinase was produced at a similar level
at different concentration of raw feathers and feather meal (Figure
4). Increased amounts of feather and feather meal resulted in higher bacterial
growth, but the cultivation with 50 g l-1 feathers
resulted in a share of fragmented quills in solution. When soy meal was used
as substrate, a decrease in keratinase activity was observed at elevated concentrations,
although this effect was less evident at longer cultivation times (Figure
4).
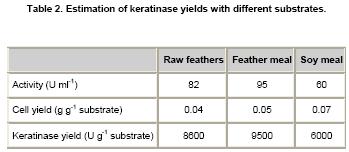
The yields for the production of keratinase using three inexpensive
common substrates were determined (Table
2). The most
effective production was achieved by using feather meal, which is 1.6 fold
higher than that in soy meal.
DISCUSSION
A keratinase was produced by Chryseobacterium sp. kr6
under different growth conditions. This enzyme has been shown to be useful
for biotechnological purposes such as hydrolysis of poultry feathers (Riffel
et al. 2003a) and de-hairing of bovine pelts (Riffel et al.
2003b).
Proteases from Chryseobacterium strains are often produced
at mesophilic temperatures. A metalloprotease of C. indologenes Ix9a
(Venter et al. 1999) and an endopeptidase of Chryseobacterium sp.
(Lijnen et al. 2000) were produced during cultivation in
nutrient broth at 25ºC and 28ºC, respectively. Although those
conditions were considered satisfactory to produce proteolytic activity, other
settings were not investigated. In the current work, biomass and enzyme activity
were found to be higher at 25-30ºC than 37ºC (Table
1).
Although keratinolytic bacteria often display optimal growth and activity at
higher temperatures (Lin et al. 1999; Kim et
al. 2001), this is consistent with optimum values described for keratinolytic
gram-negatives such as Vibrio sp. kr2 (Sangali and Brandelli,
2000), Lysobacter sp. (Allpress et al. 2002) and Stenotrophomonas sp.
D-1 (Yamamura et al. 2002), which showed optimum temperature
for growth and keratinolytic enzyme production ranging from 20ºC to 30ºC.
Maximum biomass and keratinase activity were observed from
pH 6.0 to 8.0 (Table 1), which agrees with those described
for most feather-degrading Bacillus (Wang and Shih, 1999; Suntornsukz
and Sutornsukz, 2003). For production of keratinase by B. licheniformis and
a recombinant B. subtilis, uncontrolled pH operation was more favorable
than the controlled pH operations (Wang and Shih, 1999).
The same was observed for alkaline protease production by B. licheniformis
(Çalik et al. 2002).
Maximum activities were coinciding with maximum biomass and
maximum specific production rates were observed at the exponential growth phase,
suggesting that the keratinase of strain kr6 was produced as a primary metabolite
(Figure 2).
Carbohydrate inhibition of keratinase production was observed
(Figure 3), indicating that this strain has a catabolite
repression regulatory mechanism, a common control mechanism for biosynthesis
of bacterial proteases (Givskov et al. 1991), including keratinases
(Wang and Shih, 1999; Thys et al. 2004).
Suitable amounts of some surfactants, such as Tween 80 and Triton X-100, are
known to stimulate bacterial enzyme production (Reddy et al.
1999). However, the addition of these detergents into the FKB suppressed
bacterial growth and keratinase production (Figure 3),
similar to that observed for Bacillus sp. FK46 during submerged cultivation
(Suntornsukz and Sutornsukz, 2003).
Although the production of proteases in complex growth media
often promotes exuberant growth and high enzyme yields (Johnvesly
and Naik, 2001; Joo et al. 2002), their expensive cost
makes them unsuitable for a large-scale production. Some less expensive substrates,
such as soybean meal, have been successfully used (Wang and Shih,
1999; Joo et al. 2002). It seems more adequate to use
raw materials like some wastes from the food industry as a basis of the culture
media. The strain kr6 produced higher yields in feather meal and raw feathers,
which have been used as good substrates for production of other keratinolytic
enzymes (Wang and Shih, 1999; Sangali and Brandelli,
2000).
High substrate concentrations may cause substrate inhibition
or repression of keratinase production. This was observed when soy meal was
used as substrate, similar to that described by Joo et al. (2002) during
protease production by Bacillus horikoshii. This indicates that smaller
amounts of substrate are preferred since they result in higher yields of product
per substrate (U g-1 substrate). It was previously observed for
the strain kr6 that substrate and metabolite levels in the extracellular milieu
can regulate enzyme secretion (Riffel et al. 2003a). As the
strain grows in soy meal faster than in feather substrates, different kinetics
for keratinase production would be expected, depending on the type and amount
of substrate.
Keratinases have enormous potential applications in processing
waste in the poultry and leather industries. The recent finding that B. licheniformis
PWD-1 keratinase cause enzymatic breakdown of prion protein PrPSc (Langeveld
et al. 2003) leave open a novel relevant application for broad range keratinases.
In this study, the optimum conditions for keratinase synthesis by the Chryseobacterium strain
kr6 were determined, which is an essential step for the production of adequate
amounts for application in research of feed and other areas.
REFERENCES
-
ALLPRESS, J.D.; MOUNTAIN, G. and GOWLAND,
P.C. Production, purification and characterization of an extracellular keratinase
from Lysobacter NCIMB 9497. Letters in Applied Microbiology,
May 2002, vol. 34, no. 5, p. 337-342.
-
ÇALIK, P.; BILIR, E.; ÇALIK, G. and ÖZDAMAR,
T.H. Influence of pH conditions on metabolic regulations in serine alkaline
protease production by Bacillus licheniformis. Enzyme and Microbial
Technology, October 2002, vol. 31, no. 5, p. 685-697.
-
CHOI, J.M. and NELSON, P.V. Developing
a slow-release nitrogen fertilizer from organic sources. 2. Using poultry
feathers. Journal of the American Society of Horticultural Science,
July 1996, vol. 121, no. 4, p. 634-638.
-
DALEV, P. and NEITCHEV, V. Reactivity
of alkaline proteinase to keratin and collagen containing substances. Applied
Biochemistry and Biotechnology, February 1991, vol. 27, no. 2, p. 131-138.
-
DORAN, P.M. Bioprocess Engineering
Principles. San Diego,
Academic Press, 1995. 439 p. ISBN 0-12-220856-0.
-
GIVSKOV, M.; EBERL, L. and MOLIN, S.
Control of exoenzyme production, motility and cell differentiation in Serratia
liquefaciens. FEMS Microbiology Letters, March 1991, vol. 148,
no. 2, p. 115-122.
-
JOHNVESLY, B. and NAIK, G.R. Studies on production
of thermostable alkaline protease from thermophilic and alkaliphilic Bacillus sp.
JB- 99 in a chemically defined medium. Process
Biochemistry, October 2001, vol. 37, no. 2, p. 139-144.
-
JOO, H.S.; KUMAR, C.G.; PARK, G.C.; KIM, K.T.;
PAIK, S.R. and CHANG, C.S. Optimization of the production of an extracellular
alkaline protease from Bacillus hirikoshii. Process Biochemistry,
October 2002, vol. 38, no. 2, p. 155-159.
-
KIM, J.M.; LIM, W.J. and SUH, H.J. Feather-degrading Bacillus species
from poultry waste. Process Biochemistry, November 2001, vol. 37,
no. 3, p. 287-291.
-
LANGEVELD, J.P.M.; WANG, J.J.; VAN DE WIEL,
D.F.M.; SHIH, G.C.; GARSSEN, J.; BOSSERS, A. and SHIH, J.C.H. Enzymatic degradation
of prion protein in brain stem from infected cattle and sheep. Journal
of Infectious Diseases, December 2003, vol. 188, no. 11, p. 1782-1789.
-
LIJNEN, H.R.; VAN HOEF, B.; UGWU, F.; COLLEN,
D. and ROELANTS, I. Specific hydrolysis of human plasminogen by a 24 kDa
endopeptidase from a novel Chryseobacterium sp. Biochemistry,
January 2000, vol. 39, no. 2, p. 479-488.
-
LIN, X.; INGLIS, G.D.; YANKE, L.J. and
CHENG, K.J. Selection and characterization of feather-degrading bacteria
from canola meal compost. Journal of Industrial Microbiology and Biotechnology,
August 1999, vol. 23, no. 2, p. 149-153.
-
LIN, X.; LEE, C.G.; CASALE, E.S. and SHIH,
J.C.H. Purification and characterization of a keratinase from a feather-degrading Bacillus
licheniformis strain. Applied and Environmental Microbiology,
October 1995, vol. 58, no. 10, p. 3271-3275.
-
LOWRY, O.H.; ROSEMBROUGH, N.J.;
FARR, A.L. and RANDALL, R.J. Protein measurement with the Folin phenol reagent. Journal
of Biological Chemistry, January 1951, vol. 193, no. 1, p. 267-275.
-
RAJU, A.A.; CHANDRABABU, N.K.; SAMIVELU,
N., ROSE, C. and RAO, N.M. Eco-friendly enzymatic dehairing using extracellular
proteases from a Bacillus species isolate. Journal of the American
Leather Chemical Association, May 1996, vol. 91, no. 5, p. 115-119.
-
RAO, M.B.; TANKSALE, A.M.; GHATGE,
M.S. and DESHPANDE, V.V. Molecular and biotechnological aspects of microbial
proteases. Microbiology and Molecular Biology Reviews, September 1998,
vol. 62, no. 3, p. 597-635.
-
REDDY, R.M.; REDDY, P.G. and
SEENAYYA, G. Enhanced production of thermostable β-amylase and pullanase
in the presence of surfactants by Clostridium thermosulfurogenes SV2. Process
Biochemistry, January 1999, vol. 34, no. 1, p. 87-92.
-
RIFFEL, A.; LUCAS, F.S.; HEEB, P. and BRANDELLI,
A. Characterization of a new keratinolytic bacterium that completely degrades
native feather keratin. Archives of Microbiology, April 2003a, vol.
179, no. 4, p. 258-265.
-
RIFFEL, A.; ORTOLAN, S. and BRANDELLI, A.
De-hairing activity of extracellular proteases produced by keratinolytic
bacteria. Journal of Chemical Technology and Biotechnology, August
2003b, vol. 78, no. 8, p. 855-859.
-
SANGALI, S. and BRANDELLI, A. Feather keratin
hydrolysis by a Vibrio sp. strain kr2. Journal of Applied Microbiology,
November 2000, vol. 89, no. 5, p. 735-743.
-
SINCLAIR, C.G. and CANTERO, D. Fermentation
modeling. In: McNEIL, B. and HARVEY, L.M. eds. Fermentations, a practical
approach. Oxford, IRL Press,
1990, p. 65-113. ISBN 0199630453.
-
SUNTORNSUK, W. and SUNTORNSUK, L. Feather
degradation by Bacillus sp. FK46 in submerged cultivation. Bioresource
Technology, February 2003, vol. 86, no. 3, p. 239-243.
-
THYS, R.C.S.; LUCAS, F.S.; RIFFEL, A.;
HEEB, P. and BRANDELLI, A. Characterization of a protease of a feather-degrading Microbacterium species. Letters
in Applied Microbiology, February 2004, vol. 39, no. 2, p. 181-186.
-
VENTER, H.; OSTHOFF, G. and LITTHAUER,
D. Purification and characterization of a metalloprotease from Chyseobacterium
indologenes Ix9a and determination of the amino acid specificity with
electrospray mass spectrometry. Protein Expression and Purification,
April 1999, vol. 15, no. 3, p. 282-295.
-
WANG, J.J. and SHIH, J.C.H. Fermentation
production of keratinase from Bacillus licheniformis PWD-1 and a recombinant B.
subtilis FDB-29. Journal of Industrial Microbiology and Biotechnology,
June 1999, vol. 22, no. 6, p. 608-616.
-
WANG, X. and PARSONS, C.M. Effect of
processing systems on protein quality of feather meal and hog hair meals. Poultry
Science, March 1997, vol. 76, no. 3, p. 491-496.
-
WILLIAMS, C.M.; LEE, C.G.; GARLICH, J.D.
and SHIH, J.C.H. Evaluation of a bacterial feather fermentation product,
feather-lysate, as a feed protein. Poultry Science, January 1991,
vol. 70, no. 1, p. 85-94.
-
YAMAMURA, S.; MORITA, Y.; HASAN, Q.; RAO,
S.R.; MARAKAMI,Y.; YOKOYAMA, K. and TAMIYA, E. Characterization of a new
keratin-degrading bacterium isolated from deer fur. Journal of Bioscience
and Bioengineering, June 2002, vol. 93, no. 6, p. 595-600.
Note: Electronic Journal of Biotechnology is not responsible
if on-line references cited on manuscripts are not available any more after
the date of publication.
Supported by UNESCO / MIRCEN network
© 2005 by Pontificia Universidad Católica de Valparaíso --
Chile
The following images related to this document are available:
Photo images
[ej05006f1.jpg]
[ej05006f4.jpg]
[ej05006t1.jpg]
[ej05006f3.jpg]
[ej05006t2.jpg]
[ej05006f2.jpg]
|

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}