 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Electronic Journal of Biotechnology, Vol. 8, No.2, August 15, 2005, pg. 218-225 SHORT COMMUNICATION LaNe RAGE: a new tool for genomic DNA flanking sequence
determination
Daniel J. Park Department of Zoology, The University of Melbourne, Gate 12, Royal Parade, Victoria 3010, Australia, Tel: 61 3 83444351, Fax: 61 3 83447909, E-mail: d.park@unimelb.edu.au Financial support: This work was supported by the National Health and Medical Research Council of Australia grant no. 148630 to Professors J.A.M. Graves and M.B. Renfree, Associate Professor V.R. Harley and Dr. A.J. Pask. Received July 2 , 2004 / Code Number: ej05025 Abstract The determination of genomic DNA sequence flanking a known region is often problematic. Existing technologies depend on multiple, efficient enzyme-catalysed preparative processing steps and/or rely on relatively inefficient ‘one-sided' PCR mechanisms. I demonstrate the application of a simple ‘two-sided' PCR-based approach, lariat-dependent nested PCR for rapid amplification of genomic DNA ends (LaNe RAGE), applied to the mouse GAPDH and PGK1 gene flanking sequences. This demonstration offers great promise in applications such as genome walking, transposon mutagenesis mapping and DNA fingerprinting. Keywords: cloning, DNA fingerprinting, genomic DNA walking, PCR, sequencing, transposon mutagenesis.
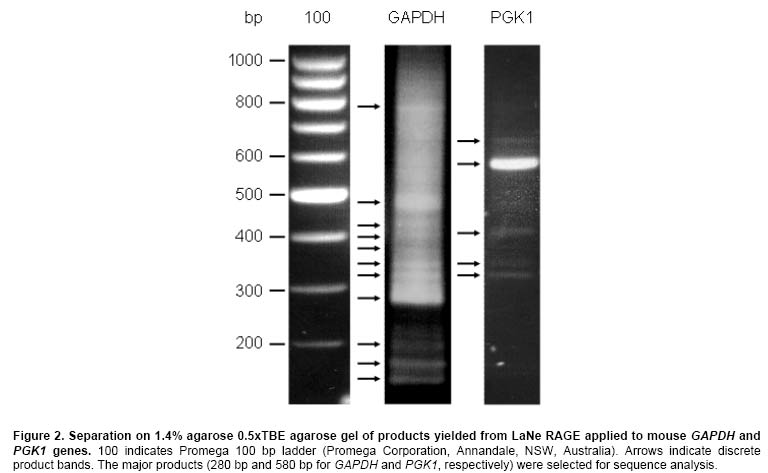
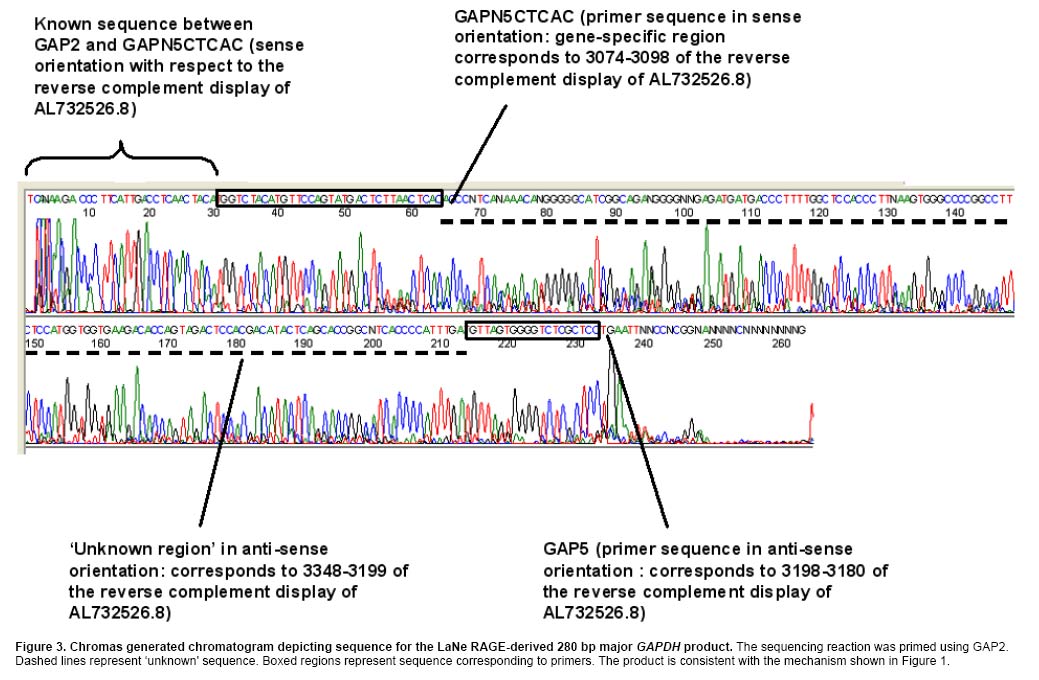
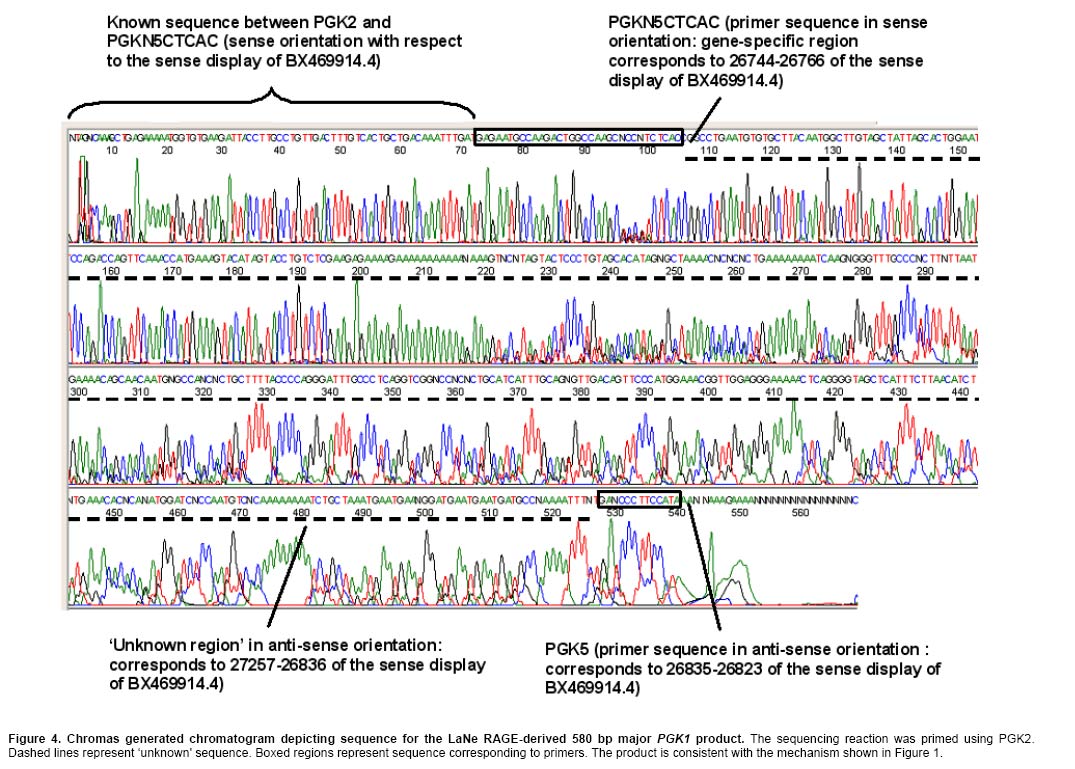
Current methods for PCR-based genomic DNA (gDNA) flanking sequence determination variously require relatively large amounts of starting material, efficient enzyme reaction steps reliant on restriction endonuclease, ligase and/or exonuclease or dependence on relatively inefficient ‘one-sided' PCR reactions (Mueller and Wold, 1989; Arnold and Hodgson, 1991; Sarkar et al. 1993; Devon et al. 1995; Myrick and Gelbart, 2002). This study set out to demonstrate the feasibility of a new technology, LaNe RAGE, requiring only limited starting material without the need for involved pre-processing steps and capable of unleashing the exquisite sensitivity of fully gene-specific ‘two-sided' nested PCR series (Figure 1). Materials and Methods gDNA was extracted from the mouse HC11 mammary gland cell line using the DNeasy Tissue Kit (Qiagen, Clifton Hill, Vic., Australia). Primers were designed based on the GenBank accession sequences NT 039711.3 (BX469914.4) and NT 039206.3 (AL732526.8 reverse complement view) for PGK1 and GAPDH, respectively. Locations within these regions used for primer design are indicated after each primer sequence. 300 ng gDNA was used as template in 50 µl PfuTurbo® DNA Polymerase (Stratagene, Integrated Sciences, East Kew, Vic., Australia) –catalysed strand synthesis reactions primed by hybrid 5' gene-specific and 3' partially defined primers PGKN5CTCAC (5'-GAGAATGCCAAGACTGGCCAAGCNNNNNCTCAC-3': 26744-26766) or GAPN5CTCAC (5'-TGGTCTACATGTTCCAGTATGACTCNNNNNCTCAC-3': 3074-3098) for mouse PGK1 or GAPDH, respectively. The semi-random 3' termini were chosen with the expectation that under the appropriate annealing conditions, they would bind at relatively frequent intervals adjacent to a known region. As such, a ladder of gene-specific products would be anticipated. The reaction mix tubes were heated to 94ºC for 3 min and then placed on ice prior to addition of 1.5 µl 10 mM dNTPs and PfuTurbo® DNA Polymerase. The reaction tubes were then heated from 18ºC to 72ºC at a rate of 0.1ºC per 5 sec and held at 72ºC for 10 min prior to placing on ice. Primers PGK1 (5'-TTGGCACATCTCTGTATGATGAAG-3': 26622-26645) and PGK4 (5'-TACTGTGGCCTCTGGTATACCT-3': 26767-26788) or GAP1 (5'-ATGGTGAAGGTCGGTGTGAACG-3': 2953-2974) and GAP4 (5'-CCGAGAATGGGAAGCTTGTCATC-3': 3131-3153), respectively, were added prior to thermocycling (36 cycles of 94ºC for 1 min; 58ºC for 1 min; 72ºC for 2 min). The lariat structure formation and template generation depicted in the large boxed section of Figure 1 occurs inherently during the thermal cycling of LaNe RAGE. 1 µl product from each first round PCR was included as template in 50 µl hot-start second round PCRs employing primers PGK2 (5'-GGAGCCAAGATTGTCAAAGATCTC-3': 26648-26671) and PGK5 (5'-CTGTTCGCTTATGGAAGGGATC-3': 26814-26835) or GAP2 (5'-CAGTGGCAAAGTGGAGATTGTTGC-3': 3018-3041) and GAP5 (5'-GGAGCGAGACCCCACTAAC-3': 3180-3198), respectively, and PfuTurbo® DNA Polymerase (40 cycles of 94ºC for 1 min; 60ºC for 1 min; 72ºC for 2 min). Products were visualized by UV trans-illumination after agarose gel electrophoresis. Major PCR products were gel extracted using the QIAEX®II bulk silica DNA purification system (Qiagen) and subjected to sequencing employing ABI PRISM®BigDye™Terminators v.3.1 (Applied Biosystems, Scoresby, Vic., Australia) primed by PGK2, PGK5, GAP2 or GAP5. Electrophoresis was performed by the Wellcome Trust Sequencing Facility, a facility of Monash Institute of Reproduction and Development and Prince Henry's Institute of Medical Research, Monash Medical Centre, Clayton, Vic., Australia. Results and Discussion Figure 2 illustrates the agarose gel profiles resulting from application of LaNe RAGE to the mouse GAPDH and PGK1 genes. The 280 bp and 580 bp products for GAPDH and PGK1, respectively, as the major products, were chosen for DNA sequence analysis (Figure 3 and Figure 4). Nucleotide-nucleotide BLAST searching and Clustal W alignment analysis, indicated that they represented GAPDH and PGK1 flanking regions, respectively (Figure 5 and Figure 6). Furthermore, the presence of gene specific hybrid primer-derived sequence confirmed that the products had been yielded via a LaNe RAGE mechanism. Major bands were consistently observed in independent experiments and were confirmed to be gene-specific by DNA sequence analysis. The GAPDH profile exhibits an expected ladder of products while that for PGK1 shows a major product and relatively few minor distinct bands. It is possible that the gene-specific region of the PGKN5CTCAC primer contributed to enhance binding at a particular site, such that the resulting major product out-competed others in the ensuing PCRs. The background smear observed in the GAPDH product could be due to primer binding at more regular intervals than anticipated (for example, TCAC or CAC sites). Altered primer design and annealing conditions change product profiles to suit the requirements of the application. For example, the design of ‘less random' primers for use in the initial annealing step should result in a ladder with longer products and bigger ‘steps'. Nonetheless, the described experiments effectively serve to demonstrate the potential of LaNe RAGE. This demonstration of a new simple and effective tool for derivation of gDNA flanking sequence requiring only limited starting material has implications not only for genome walking, but also for identification of insertion sites resulting from transposon mutagenesis screens and for DNA fingerprinting applied in contexts such as pathogenic bacteria strain typing. These applications should simply require optimization of primer design and thermocycling conditions. LaNe RAGE should offer greater specificity and sensitivity than existing approaches via a quick, simple and robust mechanism. At the heart of this is the capacity for ‘two-sided' gene-specific nested PCR series without the requirement for involved enzyme pre-processing steps. References
Note: Electronic Journal of Biotechnology is not responsible if on-line references cited on manuscripts are not available any more after the date of publication. Supported by UNESCO / MIRCEN network © 2005 by Pontificia Universidad Católica de Valparaíso -- Chile The following images related to this document are available:Photo images[ej05025f4.jpg] [ej05025f6.jpg] [ej05025f3.jpg] [ej05025f2.jpg] [ej05025f5.jpg] [ej05025f1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}