 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
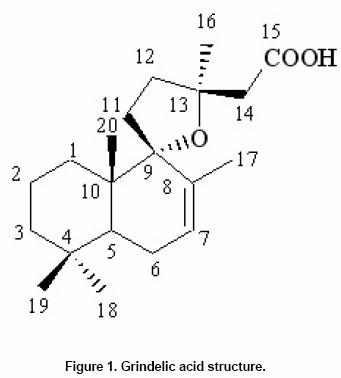
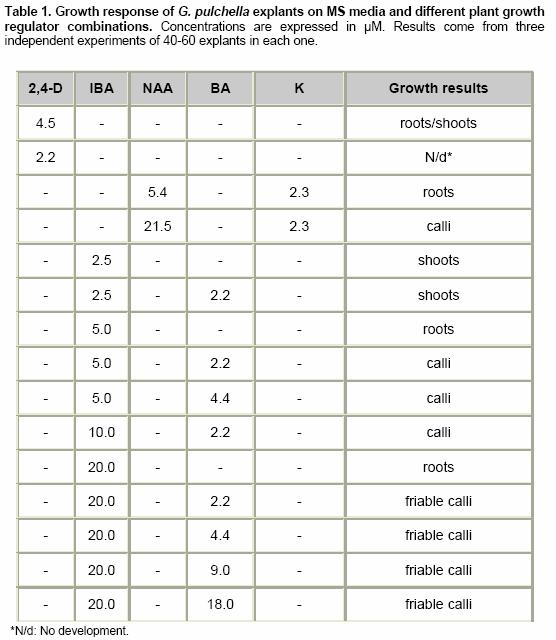
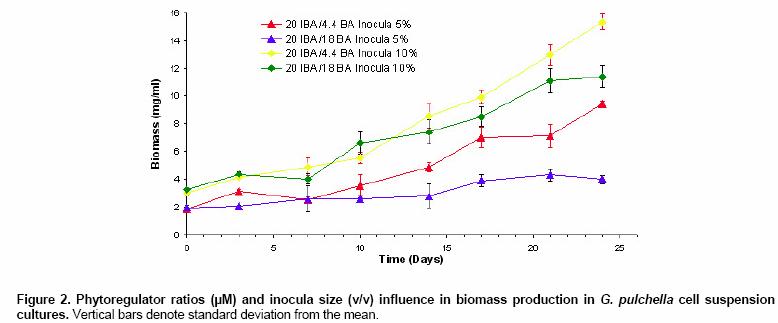
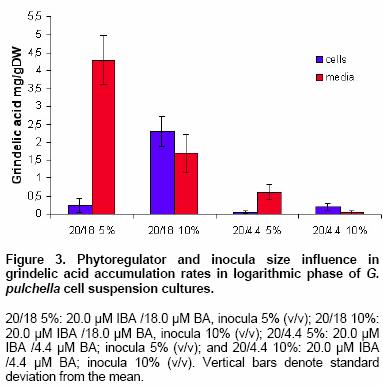
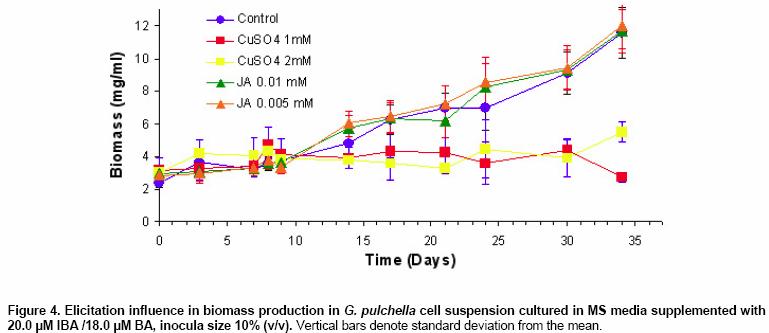
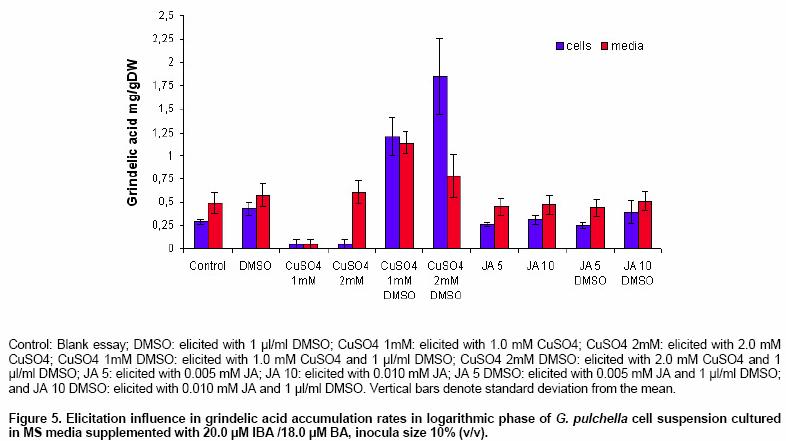
Effects of elicitor and copper sulfate on grindelic acid production in submerged cultures of Grindelia pulchella Xenia E. Hernandez1, Alejandro A. Orden2, Oscar S. Giordano3, Marcela Kurina*4 1Area de Química Orgánica (INTEQUI-CONICET), Facultad de Química, Bioquímica y Farmacia, Universidad Nacional de San Luis, Chacabuco y Pedernera, 5700, San Luis, Argentina, Tel: 54 2652439909, Fax: 54 2652426711 Financial support: UNSL: PROIPRO 0201; CONICET: PID 2429; FONCyT; ANPCyT: PICT2002 Nº 06-10710. Code Number: ej05031 Abstract Grindelia pulchella callus and cell suspension cultures were established from seedling leaves. When several phytoregulator supplementations were assayed in solid Murashige and Skoog medium containing 3% (w/v) of sucrose (MS medium), combinations of indole-3-butyric acid (IBA) and N6-benzylaminopurine (BA) resulted the most appropriate conditions to generate fast growing friable calli with detectable levels of grindelic acid. Moreover, the same basal media supplemented with 20.0 µM IBA/4.4 µM BA was found to be optimal for cell growth in submerged cultures (µmax = 0.26 days-1) while the addition of 20.0 µM IBA/18.0 µM BA resulted in a relative higher metabolite production (4.55 mg/gDW) when the inocula was 5% (v/v). Furthermore, three different stress factors and combinations of them were used to elicit cell suspensions. These experiments demonstrated that the combination of CuSO4 and dimethylsulfoxide (DMSO) increase the grindelic acid production to 2.63 mg/gDW in the elicited essay versus 0.756 mg/gDW Keywords: Cu2+, dimethylsulfoxide permeabilization, grindelane diterpenes, jasmonic acid. Abbreviations: Grindelia species are widespread in South American semiarid regions. Most of these genuses are used in folk medicine as antispasmodic and diuretic, among other purposes. Grindelic acid (Figure 1) and minoritary hydroxylated metabolites were isolated from G. pulchella aerial parts (Guerreiro et al. 1981). Grindelic acid hydroxylated derivatives, the acids 6-β-hydroxygrindelic, 6-α-hydroxygrindelic and 3-β-hydroxygrindelic, were successfully obtained by fungal bioconvertion from grindelic acid (Hernandez et al. 1997; Hernandez et al. 2002; Orden et al. 2005) and showed bioactive properties towards Tenebrio molitor larvae species and phytopatogen fungi and bacteria. Furthermore, the resins of several species of the genus Grindelia such us G. chiloensis, G. camporum and G. glutinosa, particularly rich in diterpene acid derivatives, have been extensively studied due to their possible industrial applications like pine resin and raw material for the naval stores industry (Hoffmann and McLaughlin, 1986; Timmermann et al. 1987; Ravetta et al. 1996; Ravetta and Soriano, 1998; Zavala and Ravetta, 2001; Zavala and Ravetta, 2002; Wassner and Ravetta, 2005). Even some attempts have been made to generate protocols for vegetative propagation of G. chiloensis (Wassner and Ravetta, 2000). There has been a long-standing interest in the exploration and utilization of plant cell cultures for the production of plant secondary metabolites. Searching for a biotechnological approach as an alternative for stable production of grindelic acid seems to be quite promising since a wide range of derivatives could be obtained by both chemical and biological transformations. Elicitation by heavy metals is a procedure that has demonstrated to improve secondary metabolite accumulation in plant cell cultures (Rakwal et al. 1996; Oikawa et al. 2001; Mithöfer et al. 2004). Many authors have reported evidences that jasmonates are involved in the biosynthesis of a wide spectrum of secondary metabolites (Menke et al. 1999; Vom Endt et al. 2002) and we have previously demonstrated that DMSO helps to increase terpenoid metabolite accumulation in cell cultures (Kurina et al. 2000). In a preview personal communication, we have demonstrated that grindelic acid possesses antimicrobial activity toward several phytopatogenic fungal and bacterial strains. Taking into account that the biosynthesis and accumulation of this sort of secondary metabolites may be induced by biotic elicitation, JA, a proved mediator in plant-wounding and plant-microbe interactions, was chosen to try to induce the aforementioned metabolite accumulation in vitro. The objective of this work was to establish submerged cultures of G. pulchella in order to study the possibility of producing stable amounts of grindelic acid. In this sense the effects of elicitors such as JA and copper salts were tested on DMSO-permeabilized cell suspension cultures. Materials and Methods The 1H NMR spectra were recorded in CDCl3 at 200.13 MHz and G. pulchella seeds and aerial parts were collected in Departamento Capital, Grindelic acid isolation and structure determination Grindelic acid authentic samples were isolated from aerial parts of wild G. pulchella specimens as previously reported (Guerreiro et al. 1981), and structural identity was confirmed by 1H NMR and [α] G. pulchella seeds were surface disinfected and germinated in aseptic conditions. Seedling first true leaves were used to initiate callus cultures on MS medium. Different conditions were obtained by supplementing the above-mentioned salt basal medium with different auxin/cytokinin ratios (Table 1). Incubation was carried out at 22 ± Biomass was evaluated as dry weight (DW) of cells in vacuum at Grindelic acid extraction and identification The freeze-dried calli were extracted three times with methanol in reflux. Evaporation of solvent under reduced pressure gave the methanol extract, which was then partitioned between chloroform and water. The organic phase was evaporated and subjected to preparative TLC eluted with benzene: dioxane: acetic acid (30:5:1) to give grindelic acid, which identity was confirmed by 1H NMR and Dried cells were homogenized and extracted in methanol by reflux (X3). Culture medium was acidified to pH 5/5.5 and extracted three times with Et2O. Both methanolic and ethereal extracts were washed, concentrated, and methylated with diazomethane and redisolved in acetone. Quantification of methylgrindelate was carried out by GC using an OV 17 column, a N2 flow rate of 30 ml min-1 and a temperature gradient from They were carried out on 7-day-old cultures in MS medium supplemented with 18.0 µM BA and 20.0 µM IBA. DMSO and CuSO4 solutions were sterilized by filtration. DMSO was added to the cultures at final concentrations of 1.0 µl per ml of culture. Meanwhile CuSO4 solutions were added to reach a final concentration of 1.0 and Results and Discussion G. pulchella in vitro culture establishment Calli of G. pulchella could be successfully induced from in vitro germinated seedlings when explants from different organs were cultured on MS media added with several plant growth regulator combinations and ratios (Table 1). The best results for callus production were obtained when explants were cultured in the presence of 20.0 µM of IBA and 18.0 µM of BA. Other auxin/cytokinin combinations and ratios were less effective in promoting callus development. Although several authors demonstrated that 2,4-D was necessary to initiate callus induction (Banthorpe, 1994), it was ineffective in our experiences and, even necrosis was evident after the first subculture. Microscopy revealed the presence of adult cells organized in differentiated tissues, particularly vascular ones. NAA tended to induce the regeneration of roots, independently of the origin of the explants. The presence of grindelane diterpenes in the different callus lines was checked by extraction and further TLC analysis compared with authentic samples. Grindelic acid was detected only in calli grown in MS media supplemented with different IBA/BA combinations. Its identity was confirmed by 1HNMR and 13CNMR spectroscopy. No other minor grindelane metabolites were observed by this methodology in the analyzed biomass samples. Cell suspension cultures and grindelic acid accumulation Suspension cultures were initiated from friable tissues using those callus lines developed in MS basal media supplemented with 20.0 µM IBA/18.0 µM BA and 20.0 µM IBA/4.4 µM BA that showed detectable grindelic acid accumulation as it was described above. Significant differences were observed when cell growth ratios of cultures grown in basal media supplemented with 20.0 µM IBA/18.0 µM BA were compared with those added with 20.0 µM IBA/4.4 µM BA using inocula sizes of 5 and 10 % v/v in both cases. In the former phytoregulator condition, a µmax = 0.177 days-1 was obtained when inocula was of 10%, and µmax = 0.052 days-1 with inocula of 5%. However, when MS media was supplemented with 20.0 µM IBA/4.4 µM BA, µmax = 0.23 days-1 and µmax = 0.26 days-1 for inocula of 10% and 5%, respectively, were observed (Figure 2). Intra and extracellular grindelic acid accumulations were evaluated. The maximal grindelic acid levels during logarithmic growth phase were registered. Nevertheless, no other grindelane metabolites were detected neither in biomass nor in culture media. Unfortunately, the best culture conditions for cell growth showed the minimal grindelic acid accumulation rates. Interestingly, grindelic acid production increased and it was a noticeable positive effect on grindelic acid extracellular accumulation when biomass inoculum was diminished to 5%. Since the rates of extracellular accumulation increased as the intracellular quantities were reduced, it seems that grindelic acid was excreted to the media more efficiently in this condition than in more dense cultures (Figure 3). Elicitation effects on cell growth 15 monthly subcultures were performed in order to obtain stable cell lines. After this period, not only grindelic acid accumulation rates drastically diminished, but also intra and extracellular ratios changed. These results might have been caused by epigenetic changes or epimutations (Martienssen and Colot, 2001) caused by cell dediferentiation process that often lead to a lower productivity (Verpoorte et al. 2002). It has been reported that octadecanoid elicitors affected isoprene metabolite biosynthesis and accumulation in a number of plant species. Namely, the interaction of jasmonates with wounding and/or fungal elicitation in Hyoscyamus muticus root cultures (Singh et al. 1998); in Solanum tuberosum (Choi et al. 1994) and in elicited cell suspension of Tessaria absinthioides (Kurina et al. 2000). Moreover, in the last specie, when the solvent DMSO was added as a permeabilizing agent, it acted as an abiotic elicitor (Kurina and Donadel, 2003). When DMSO was used to permeabilize the G. pulchella cell suspension cultures maintained in MS basal media amended with 20.0 µM IBA/18.0 µM BA, cell growth rates did not show statistical significant differences with the controls. Furthermore, the growth of G. pulchella cell suspension was not affected by the addition of JA although it was drastically suppressed by CuSO4 treatments as it is shown in Figure 4. Elicitation effects on grindelic acid accumulation Grindelic acid contents were determined during logarithmic growth phase after the elicitor addition. Highest grindelic acid accumulations in biomass and culture media were expressed as mg of grindelic acid per cell dry weight and are shown in Figure 5. Surprisingly permeabilization experiments did not result in a statistical significant increase in the grindelic acid rates excreted from cells to media. Furthermore, a slight elicitation effect was observed in cultures treated only with DMSO. Grindelic acid rates were higher in the controls that in the cultures elicited with 1.0 and Concluding Remarks This work results in a valuable contribution to establish in vitro culture conditions for the wild specie G. pulchella in order to produce the diterpene grindelic acid. In this sense different IBA/BA combinations resulted in the best phytoregulator supplementation to establish G. pulchella callus and cell suspension cultures. Meanwhile the addition of 20.0 µM IBA/4.4 µM BA was found to be optimal for biomass development in submerged culture, MS media with 20.0 µM IBA/18.0 µM BA resulted in a relative higher grindelic acid production. The elicitation experiments demonstrated that the association of CuSO4 and DMSO increases the terpenoid compound production at expense of cell growth. In contrast, neither cell growth nor grindelic acid accumulation was affected with the addition of JA alone and combined with DMSO. Further elicitation experiments such as the use of biotic agents and combinations of biotic and abiotic elicitors would allow improving grindelic acid production. According to these results, other mediators different of JA should be involved in the transduction of the elicitor signals in the regulation of the expression of diterpenoid metabolites in this system. Acknowledgments Thanks are due to Fabricio Pena for the statistical analyses and Lic. Mónica Ferrari for the technical assistance. This work is part of X.E.H.'s Doctoral thesis and has been partially presented as a poster communication at the XII National Symposium of Organic Chemistry: XII SINAQO (Hernandez et al. 2000). References
© 2005 by Pontificia Universidad Católica de Valparaíso -- Chile |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}