 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Electronic Journal of Biotechnology, Vol. 9, No.1, January 15, 2006, pg. 26-39 RESEARCH ARTICLE Citric acid production from glucose by yeast Candida oleophila ATCC 20177 under batch, continuous and repeated batch cultivation Savas Anastassiadis*#1, Hans-Jürgen Rehm2 1Research
in Biotechnology, Co.,
Vat. #: 108851559. Avgi/Sohos,
57002 Thessaloniki, Greece,
Tel: 30 2395 051324,
Fax: 30 2395 051470,
E-mail: sanastassiadis@netscape.net Financial support: The work has been carried out at the Institute of Biotechnology 2 of Research Centre Jülich (Germany) and was financed by Haarmann and Reimer, a daughter company of the company Bayer, Leverkusen, Germany. Received November 22, 2004 / Accepted May 31, 2005 Code Number: ej06004 Abstract The effect of air saturation on citric acid production in batch, repeated batch and chemostat cultures has been studied. It was shown that, under continuous fermentation (chemostat mode), the highest concentration of citric acid equal of 98 g/l was produced at 20% of air saturation. In contrary to continuous fermentation, displaying an optimum at 20%, 80% air saturation yielded higher values in repeated batch fermentation process. 167 g/l citric acid were produced continuously with the fill and drain technique at 4.85 days, at 80% air saturation, compared with 157.6 g/l achieved within 5.4 days at 20%. Under repeated batch fermentation, the formation rate of the generic product (Rj) as well as the specific citric acid productivity (mp) reached a maximum of 1.283 g/(l x hr) at 4.01 days and of 0.0375 g/(g x hr) at 4.58 days, respectively. The glucose consumption rate (Rx) reached a maximum value of 3.33 g/(l x hr) entering stationary phase after 2.56 days at a glucose concentration of 131.2 g/l. Keywords: air saturation influence, chemostat, citric acid, citric acid fermentation, repeated batch, oxygen influence.
Citric acid is an important multifunctional organic acid with a broad range of versatile uses in household and industrial applications that has been produced industrially since the beginning of 20th century. Several hundreds of thousands metric tons of citric acid are produced worldwide every year almost exclusively by fermentation. Although processes involving yeasts of genera Candida have been well described, which can produce large amounts of citric acid, industry still employs selected strains of Aspergillus niger. However, yeast processes are also sporadically used in industry today. Formation of citric and isocitric acid by yeasts using various C-sources has been the focus of investigations in different aspects for the past 35 years. Most reports are devoted to various types of citric acid producing yeasts and to experimental conditions governing production of citric acid, essentially based on shake flasks assays under undefined fermentation conditions. Only a few optimisation studies have been carried out in stirred fermenters. Continuous operation for the production of citric acid by yeasts, using glucose or other carbon sources, have increasingly received research interest in the last years; however the yields achieved were not high enough for a competitive economic industrial operation (Enzminger and Asenjo, 1986; Kim et al. 1987; Klasson et al. 1989; Kim and Roberts, 1991; Bubbico et al. 1996; Arzumanov et al. 2000; Anastassiadis et al. 2005; Anastassiadis and Rehm, 2005). Yarrowia
lipolytica and other Candida species are capable of producing
citric acid using various carbon sources and substrates (Hattori
et al. 1974; Ilarionowa et al. 1975; Furukawa
et al. 1977; Behrens et al. 1978; Lozinov
and Finogenova, 1982; Stottmeister et al. 1986; Stottmeister
and Hoppe, 1991; Anastassiadis et al. 1993; Anastassiadis,
1994; Anastassiadis et al. 1994; Moresi,
1994; Mayilvahanan et al. 1996; Tisnadjaja
et al. 1996; Anastassiadis et al. 2001; Kamzolova
et al. 2003; Anastassiadis et al. 2005), whereby
glucose has increasingly received interest after the oil crisis of
Optima
air concentration of more than 50% and up to 60 ppm are reported abroad
in international literature for discontinuous citric acid processes utilizing
yeast strains (Stottmeister et al. 1981; Okoshi
et al. 1987). Increasing aeration intensity and agitation rate has
been reported to influence positively citric acid production in various
yeast strains utilizing n-paraffin as carbon source (Hattori
et al. 1974; Ilarionowa et al. 1975; Furukawa
et al. 1977). Marchal et al. (1977) didn’t find any
further increase of citrate production rate in Saccharomycopsis lipolytica at
air saturations higher than 40% utilizing n-paraffins. An affinity
constant for oxygen of Ks=0.8 x 10-
The object of the present work is to examine the air saturation influence on repeated batch fermentation of citric acid and to compare it with continuous and batch fermentation. A new developed fermentation process is presented as an alternative innovative approach that appears suitable and commercially valid for continuous industrial production of citric acid by yeasts. Candida
oleophila ATCC 20177 var. (obtained from Dr. Siebert, Jungbunzlauer
Co. and later Haarmann and Reimer, a daughter company of Bayer Co., Leverkusen,
Germany) was used through all of experiments in present work, which was
selected under many yeast strains during an extensive screening program
(Anastassiadis et al. 1993; Anastassiadis,
1994; Anastassiadis et al. 1994; Anastassiadis
et al. 2001; Anastassiadis et al. 2002; Anastassiadis
et al. 2005). Yeast malt extract agar plates (YME) inoculated with C.
oleophila were incubated for 2-3 days and stored at
Cells
were grown in a Batch and repeated batch fermentations The
repeated batch fermentation is an alternative form of operation for continuous
citric acid production. At the end of batch fermentation a part of fermentation
solution was kept in fermenter as starting inoculum (about 300 ml corresponding
to about 1/7 of Culture conditions for batch and repeated batch experiments An extensively
optimised fermentation medium of following composition was used during
this work for batch and repeated batch experiments: 6 g/l NH4Cl,
400 g/l glucose, 1.05 g/l KH2PO4, 1.4788 g/l MgSO4 x
7H2O, 0.33435 g/l MnSO4 x 4H2O, 0 g/l
FeSO4 x 7H2O, 4 mg/l CuSO4 x 5H2O,
0.0839 g/l ZnSO4 x 7H2O, 8 mg/l CoSO4 x
7H2O, 0.08 g/l H3BO3, 0.2 g/l CaCl2,
0.2 g/l NaCl, 0.2 mg/l KJ, 0.4 mg/l Na2MoO4 x 2H2O,
2.5 g/l citric acid, 4 mg/l Thiamine-HCl, 0.5 mg/l Biotin, 1.25 mg/l Pyridoxine-HCl,
1.25 mg/l Ca-D-Pantothenate, 1 mg/l Nicotinic acid. Vitamins and NH4Cl
were added separately into autoclaved medium (sterilized for 30-60 min
at The air saturation (%) was measured using a galvanic electrode (type Ingold) and pO2 instrument (type O2-amplifier, Endress & Hauser GmbH). Air saturation was on line monitored and values were transferred to a computer program (MEAS, Research Centre Jülich, Germany). Air
saturation was adjusted to 100% using atmospheric air at 1 vvm flow rate,
1000 rpm agitation, Optical
density (OD660 nm) and dry biomass (filter method) were measured
as described in Anastassiadis et al. 2002. Organic acids,
glucose, ammonia nitrogen and intracellular concentrations were analysed
as described in Anastassiadis, 1993; Anastassiadis
et al. 1993; Anastassiadis et al. 1994; Anastassiadis
et al. 2001; Anastassiadis et al. 2002; Anastassiadis
et al. 2005. HPLC methods were developed in order to achieve a separation
between the chemically related compound citric acid and isocitric acid
or glucose and gluconic acid (Anastassiadis, 1993, Anastassiadis
et al. 1993; Anastassiadis et al. 1994; Anastassiadis
et al. 1999; Anastassiadis et al. 2001). A solvent
with
Oxygen influence on continuous citric acid fermentation by Candida oleophila ATTC 20177 (var.) During
an extensive chemostat optimisation research program, continuous citric
acid fermentation by C. oleophila,carried out applying an optimised
fermentation medium and optimum temperature of
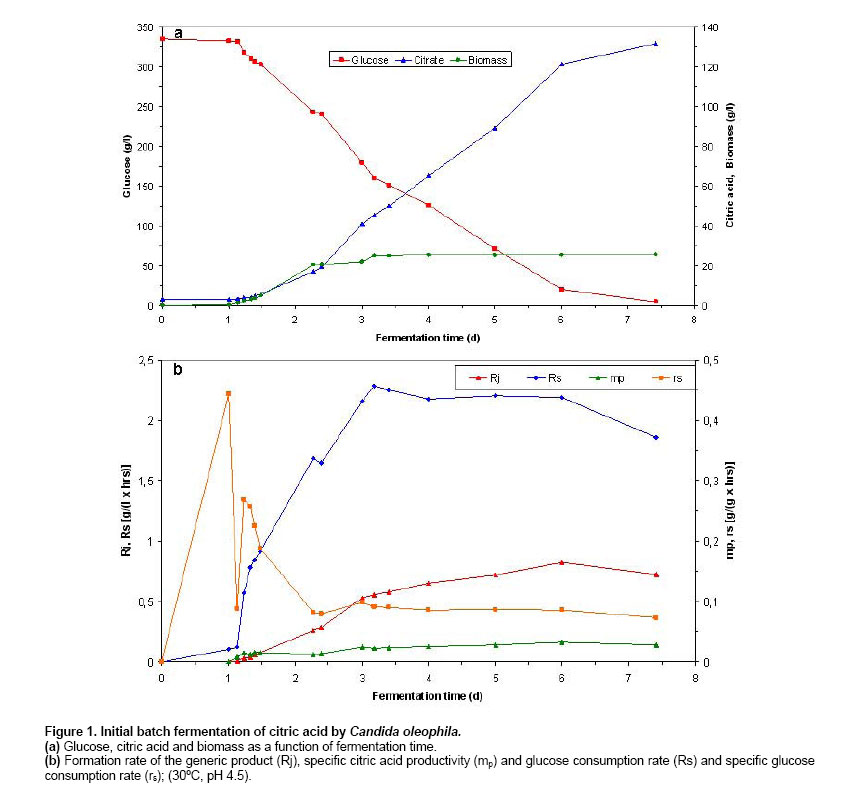
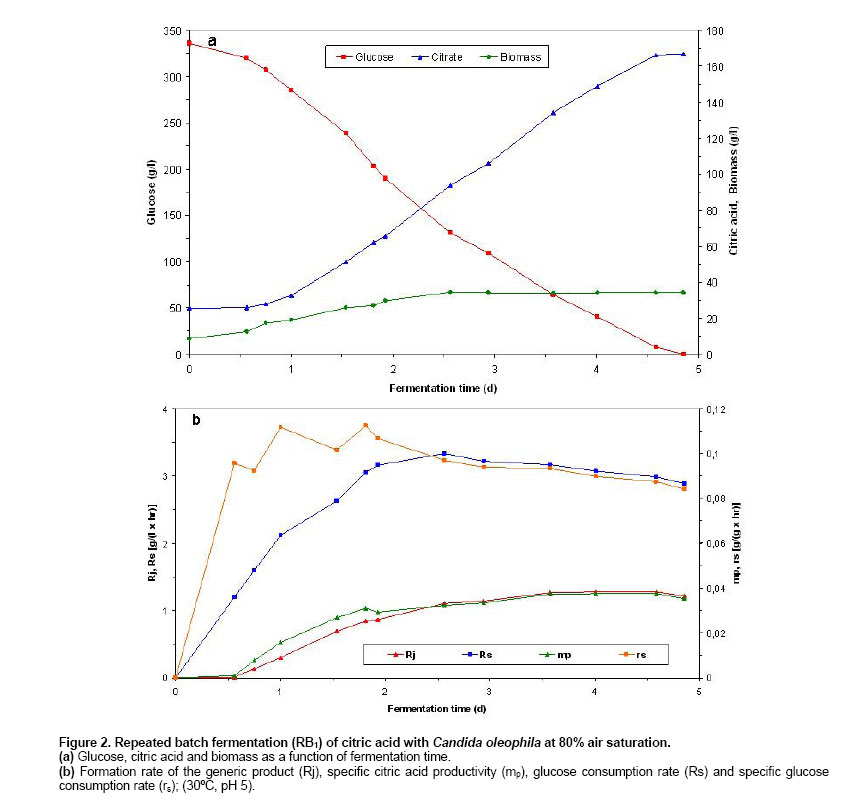
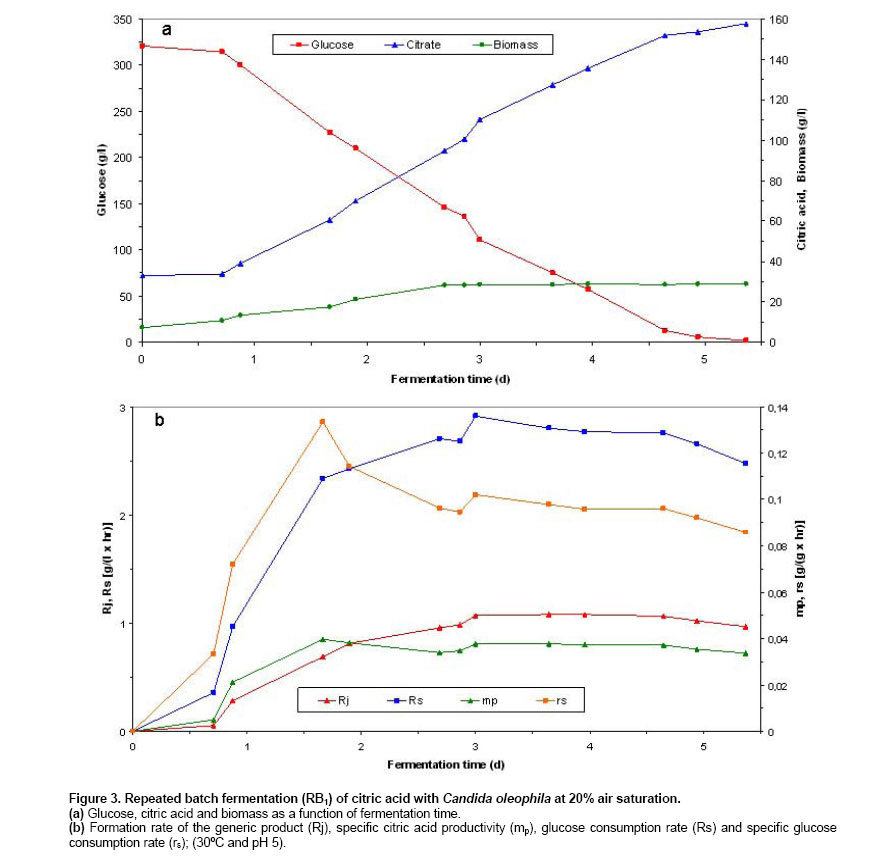
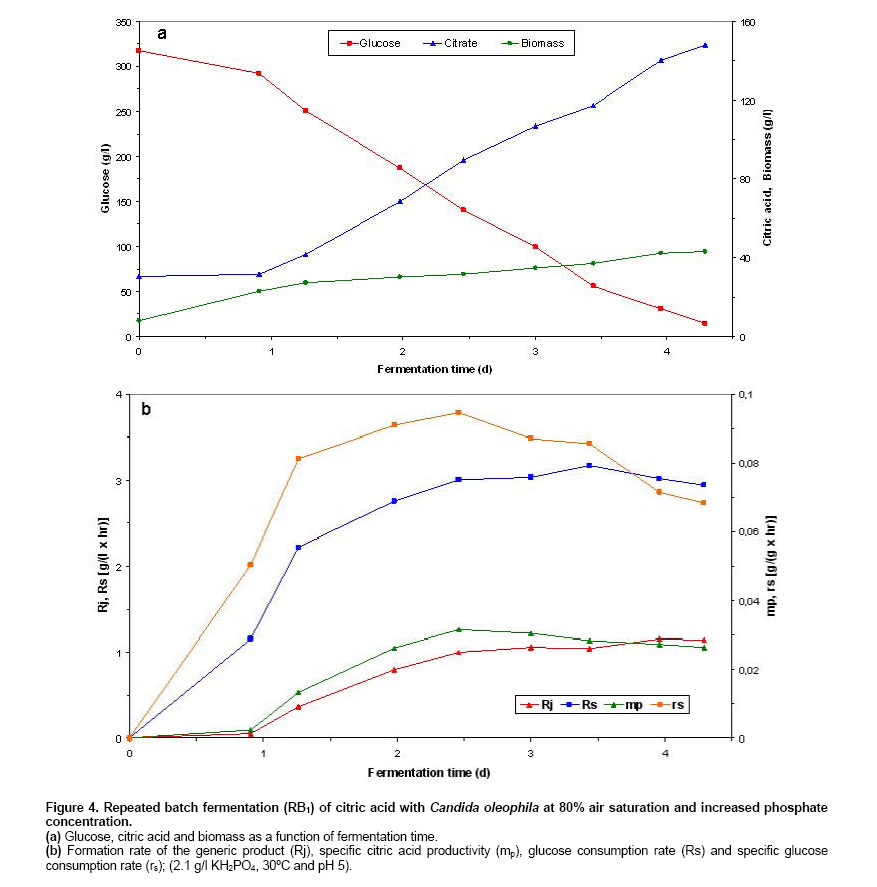
Batch and continuous repeated batch fermentation of citric acid by Candida oleophila ATTC 20177 (var.) Initialisation batch citric acid fermentation by C. oleophila ATCC 20177 (var.). Initially, the first batch fermentation was inoculated with a 10% inoculum of a 2 day culture (grown on basal medium at 30oC and 200 rpm) and it was carried out at 20% air saturation. As Figure 1 shows, starting with a biomass of 0.038 g/l and after a lag phase of about one day, biomass increased logarithmically until reaching 20.33 g/l at 2.27 days and 17.08 g/l citric acid. A biomass of 25.47 g/l, 134.3 g/l citric acid and 4.4 g/l glucose were obtained at fermentation end after 7.4 days, with a ratio between citrate and isocitrate of 15.8 and a selectivity of above 40%. Citric acid secretion started after about 1.14 days showing a steeper coarse entering post-logarithmic growth (2.27 days) and during transition (higher production) (2.27-3.19 days) and stationary growth phase (3.19-6 days), holding as long as 6 days, reaching 121.33 g/l citric acid and 20.2 g/l residual glucose. The initiation of citric acid secretion started a few hours after the complete depletion of nitrogen in the medium and was caused by the intracellular nitrogen limitation (following a transition phase) and the reduction of cellular nitrogen content that resulted in intracellular NH4+ accumulation as has been described in Anastassiadis et al. 2002. The integral formation rate (calculated from measurements of each particular point and from measurement at time zero) of the generic product (Rj) and specific citric acid productivity (mp) reached highest values of 0.82 g/(l x hr) and 0.0324 g/(g x hr) at 6 days fermentation time. A decrease in glucose consumption rate (Rs) occurred each time, sequentially entering into the next growth phase, e.g. transition phase (from 1.68 to 1.64 g/(l x hr) at 2.369 days) or stationary phase (from 2.285 to 2.253 g/(l x hr) at 3.41 days). Finally, Rs reached 1.86 g/(l x hr) at the end of fermentation after 7.42 days. The specific glucose consumption rate reached a maximum of 0.268 g/(g x hr) at 1.23 days (Figure 1). Repeated batch fermentation at 80% air saturation (RB1). Repeated batch fermentations were carried out at pH 5 and 80% (RB1) or pH 5 and 20% air saturation (RB2) in order to investigate the influence of air saturation on batch citric acid production as a comparison with the continuous chemostat fermentation. Acceleration in biomass and citrate production was observed in all of repeated batch experiments, compared with the initial batch fermentations. Figure 2 shows biomass and citric acid concentration at 80% air saturation (RB1). Despite the very initial high glucose concentration of 336 g/l, the biomass increased after a very short lag phase, indicating the very high osmotolerance of yeast strains. A biomass concentration of 29.6 g/l was determined after 1.93 days. Citric acid secretion started after about 0.56 days compared with more than 1 day in batch fermentation and was also excreted during the logarithmic growth. Following biomass formation, citric acid concentration increased continuously in a linear mode starting after a lag phase and the initiation of citric acid secretion at between 0.5 and 0.6 days, holding as long as glucose achieved a concentration of around 70 g/l. Fermentation started with an initial citric acid concentration of 25.51 g/l and biomass concentration of 8.72 g/l from previous repeated batch experiment and reached 166.5 g/l and 34.28 g/l after 4.58 days with a citrate/isocitrate ratio of 20 at a residual glucose concentration of 7.6 g/l, whilst 93.7 g/l citric acid were already measured after 2.6 days of fermentation. Glucose was finally completely consumed at last measured point of 4.85 days. Thus, a time reduction of about 2 days was achieved, compared with the batch experiment. The formation rate of the generic product (Rj) as well as the specific citric acid productivity (mp) showed a continuing increase following accumulation of citric acid, until reaching a maximum of 1.283 g/(l x hr) at 4.01 days and of 0.0375 g/(g x hr) at 4.58 days, respectively. Rs reached highest value of 3.33 g/(l x hr) entering stationary phase after 2.56 days at a glucose concentration of 131.2 g/l. The specific glucose consumption rate reached a maximum of 0.112 g/(g x hr) at 1.81 days (Figure 2). Repeated batch fermentation at 20% air saturation (RB2). As Figure 3 shows, in repeated batch experiment with 20% air saturation (RB2) the biomass followed a similar course to 80% entering stationary phase after 2.69-2.86 days. A biomass of 28.7 g/l was determined after 4.65 days. A final biomass concentration of 28.82 g/l was determined after 5.36 days, starting with a biomass of about 7.2 g/l (Figure 3). 20% air saturation resulted to lower productivity and concentration of citric acid, compared with 80%. Citric acid secretion started after about 0.71 days (33.9 g/l) and only 153.7 g/l citric acid were obtained after 4.94 days with a citrate/isocitrate ratio of 19.2, starting with an initial concentration of 33 g/l and reaching a final concentration of 157.58 g/l at 5.36 days and a residual glucose of 1.9 g/l, starting with an initial glucose concentration of 336 g/l (Figure 3). The highest Rj of 2.91 g/(l x hr) and mp of 0.102 g/(g x hr) were found after the beginning of stationary phase at about 3 days. Rs showed, with exception of a decrease from 2.71 to 2.68 g/(l x hr) at 2.86 days, a regular continuous increasing tendency until reaching highest value of 2.91 g/(l x hr) after entering stationary phase at about 3 days and 145.7 g/l glucose. Thereafter, Rs decreased continuously achieving 2.47 g/(l x hr) at the end of fermentation after 5.36 days. The highest specific glucose consumption rate (rs) of 0.134 g/(g x hr) was found at 1.67 days (Figure 3). Repeated batch fermentation at 80% air saturation and increased phosphate concentration (RB3). In a further RB experiment (RB3), performed at pH 5 and 80% saturation, the concentration of KH2PO4 was increased from 1.4 g/l to 2.1 g/l, based on medium optimisation carried out in chemostat experiments, in order to examine phosphate influence in relation to active transport system for citrate secretion and energy state of cells. Biomass increased enormously (20.5%), just changing the concentration of only this medium compound, however no significant changes were observed in citrate productivity whereas final mp decreased substantially by about 28.5%. A biomass of 43.14 g/l, 147.9 g/l citric acid and 14.5 g/l residual glucose were determined in RB3 after 4.29 days (Figure 4). The highest Rs of 3.17 g/(l x hr) was found at 3.43 days and highest mp of 0.315 g/(g x hr) was determined at 2.46 days (Figure 4). A new process for the production of citric acid in a continuous mode has been developed and presented here, that uses the filling and drain method. The experiments were repeated as long until the results achieved stable and reproducible values (a kind of steady state situation), reflecting the real situation of a series of sequential repeated batch experiments. The acceleration of fermentation can be explained by the higher starting biomass concentration in fermenter and by the adaptation of biomass to very high osmotic pressures (e.g. very high glucose or citric acid concentration). It seems very possible, that the biomass that sticks on the fermenter glass vessel can also influence the starting situation of fermentation, because it is difficult to avoid its entrance into fermenter broth during the filling procedure. In a series of repeated batch experiments, the very high citric acid concentrations were repeatedly achieved for about 20 times. No reduction had been observed in productivity, caused by the aging of cells, and the decrease in biosynthetic activity of cells, during the long cultivation time in repeated batch fermentations. The cells are revived any time the fermentation vessel is filled with a new fermentation medium. In contrary to chemostat fermentations, growth and production are running independent from each other in batch operations. In batch process, a characteristic series of events is occurring during citric acid fermentation in yeasts, starting with the extracellular depletion of nitrogen (trophophase), followed up by a transition phase of a few hours (Stottmeister et al. 1986; Behrens et al. 1978, 1987; Anastassiadis 1994; Anastassiadis et al. 2002), which is characterised by the intracellular nitrogen limitation and the reduced nitrogen content in the cells (Briffaud and Engasser, 1979; Anastassiadis et al. 1993; Anastassiadis 1994; Moresi, 1994; Anastassiadis et al. 2002). The raising intracellular NH4+ concentration (Anastassiadis et al. 2002), the increase of energy charge (Kleber and Schlee, 1987; Anastassiadis et al. 2005), cell differentiation and the vacuolisation of cells follow (Kozlova et al. 1981; Anastassiadis et al. 2002). Citric acid secretion starts a few hours after the complete depletion of nitrogen and is caused by the intracellular nitrogen limitation. In chemostat cultures of Candida oleophila, citric acid production is occurring under nitrogen limiting steady state conditions and at decreased cell nitrogen content as well (Anastassiadis et al. 2005). The active pH dependent transport system and citrate secretion by the specific active transport system are induced entering the idiophase (Anastassiadis et al. 1993; Anastassiadis 1994; Anastassiadis et al. 1994; Anastassiadis et al. 2001; Anastassiadis et al. 2002; Anastassiadis et al. 2005). Furthermore, respiration rate has been reported to decrease when nitrogen source becomes completely exhausted (Wentworth and Cooper, 1996). The intracellular nitrogen limitation has been shown to be the most important factor that triggers out citric acid secretion, although high intracellular citric and isocitric concentrations occurred under non production conditions, before the induction of secretion by a specific active transport system (Anastassiadis et al. 2002). The iron effect on continuous citric acid fermentation was optimized in details and will be published in a separate paper. Increasing iron concentration promoted biomass formation instead of citrate secretion. The best results were obtained without any addition of iron into fermentation medium. Air saturation has been found to have a very strong influence on continuous, batch and repeated batch fermentation of citric acid, indicating the importance of oxygen demand and oxygen mass transfer. The oxygen mass transfer is connected with Kla-value, which means the volumetric mass transfer coefficient of oxygen and which is strongly influenced by the viscosity of fermentation broth, aeration rate, agitation rate, temperature, the size of air bubbles etc. Most of published works in literature are carried out at unknown air saturation or dissolved oxygen concentration. In present work the air saturation was controlled, enabling the reproduction of results in future studies. The effect of air saturation was significant, resulting in differences regarding citric acid productivity of more than 20%, which would influence the costs of an industrial fermentation process enormously. Moreover, the product yield and product selectivity are also strongly influenced by the air saturation. In the case of chemostat cultures, a very high glycolytic flow rate was achieved at 20% air saturation, meaning that a kind of a Crabtree effect was attained in this case, simulating an anaerobic glycolytic pathway under aerobic conditions. The formation rate of the generic product strongly coincided with the ratio between ATP and ADP at varying the pH. The excessive energy gained by the substrate phosphorylation (high energy charge) is used promoting the active secretion of citric acid by the pH dependent active transport system (Anastassiadis and Rehm, 2005). Citric acid excretion, which starts after the exhaustion of nitrogen source and the intracellular nitrogen limitation, is promoted in repeated batch experiments in case of 80% air saturation, assuming a higher ATP level (low AMP content in cell) that has been found to closely correspond to stronger citric acid excretion by specific active transport system, found at optimum pH in C. oleophila (Anastassiadis et al. 2005; Anastassiadis and Rehm 2005). Citric acid fermentation by yeasts as well as fungi is an aerobic process, strongly depending on oxygen supply and air saturation in fermenter. An excess of oxygen is required during the entire citric acid fermentation, whereby even short time interruptions of oxygen supply can cause irreversible changes in production ability or complete production lost (Rehm, 1980; Stottmeister and Hoppe, 1991; Sakurai et al. 1996). In contrary to continuous citric acid fermentation by C. oleophila displaying an optimum air saturation of 20%, 80% saturation has been found to result to higher productivity and selectivity in batch as well as repeated batch process instead (present work). A strong decrease in specific activities was found at air saturations lower or higher than 20%, and a mp almost twice times higher was determined at 20% saturation (lower biomass concentration) compared with 80% (Anastassiadis et al. 1993; Anastassiadis, 1994; Anastassiadis et al. 1994). The very high citrate/isocitrate ratio that was achieved by C. oleophila through a sophisticated process and medium optimisation is comparable with those that have been reached by high ratio mutant yeast strains (Akiyama et al. 1972; Akiyama et al. 1973a; Akiyama et al. 1973b). Alternatively, 167 g/l citric acid were reached in continuous mode of repeated batch fermentations with the fill and drain technique after about 4.85 days at 80% air saturation, compared with only 157.6 g/l, which were achieved after about 5.4 days at 20%. In every repeated batch run, cell growth and citric acid secretion started with a shorter lag-phase compared with batch operations, reducing enormously the total fermentation time. The lag phase is strongly influenced by the osmotic pressure at the very high glucose concentrations higher than 350 g/l, which would strongly inhibit the growth of most of microorganisms. Increase of phosphate concentration resulted only in a higher biomass concentration. Wentworth and Cooper (1996) reported about the advantages of synchronous cell growth in an efficient self-cycling two-stage fed-batch fermentation process, eliminating down times that are necessary for cleaning, sterilization and inoculation, and enhancing biomass formation rate, in comparison with conventional non-synchronous growth. There is a wide distribution of cell ages and only a fraction of the cells is being at the appropriate production state at the initiation of second fermentation phase. A maximum concentration of 11 g/l was reached in shake flasks for an initial glucose concentration of 50 g/l in the second fermentation stage (Wentworth and Cooper, 1996).Tisnadjaja et al. 1996 reported about beneficial cell immobilization of C. guilliermondii from the viewpoint of recovery and reuse of cells in repeated batch cultures, reaching similar citric acid concentration, productivity and citrate yield, compared with batch cultures utilizing freely suspended cells. A little information can be found in literature regarding continuous citric acid production, in especially for A. niger process. Because for long time it has been thought that citric acid couldn’t be produced in a continuous mode, this type of data for microbial growth and continuous production of citric acid on glucose or other carbon sources is rather rare in literature. For instance, looking in the internet about 15,200 results was found as compared with 89 for chemostat (Anastassiadis et al. 2005; Anastassiadis and Rehm, 2005). Previous studies have demonstrated the feasibility of a semi-continuous production of citric acid by Aspergillus niger (Lesniak and Stawicki, 1979) and low-rate continuous production of citric acid under nitrogen-limiting conditions by yeast Candida lipolytica (Aiba and Matsuoka, 1979; Kim et al. 1987; Klasson et al. 1989). Kristiansen and Sinclair, 1979 reported about a continuous single-stage citric acid fermentation under nitrogen limitation using A. foetidus, reaching citric acid concentrations up to 40 g/l at dilution rates between 0.04 and 0.21 h-1 (4.76-25 hrs RT) and a maximum productivity of 0.43 g/(l x hr) at a dilution rate of 0.075 h-1 (13.3 hrs RT). Wieczorek and Brauer, 1998 reported about a four stage process for the continuous citric acid production using A. niger, recirculating fermentation broth, that includes the fermentation, extraction, reextraction and the product stage. A considerable increase in productivity in a continuous single-stage process using A. niger, compared with the batch process, has also been reported by Kristiansen and Charley, 1981, however only 26.5 g/l of citric acid were produced at a RT of 13 hrs and pH 1.7. In comparison, about 125 g/l citric acid were continuously produced by C. oleophila in chemostat cultures at 54 hrs RT, up to 150 g/l citric acid at higher RT and about 170 g/l in repeated batch experiments (Anastassiadis et al. 1993; Anastassiadis, 1994; Anastassiadis et al. 1994; Anastassiadis et al. 2001). Table
2 compares the new developed processes with literature reports.
Details of fermentation processes used by companies are closely guarded
and hold as trade secrets and therefore a little information is available
regarding processes used for large-scale production of citric acid.
About 150-180 g/l citric acid are commonly produced discontinuously
in industry within 5-6 days using high selected Aspergillus niger mutant
strains of several generations, which also require additional down
times for the induction of sporulation, sporulation, spore cultivation
in a smaller fermenter, preparation of fermenter and finally inoculation
of large production fermenter. The values of citric acid concentration
and formation rate of the generic product that have been obtained in
continuous mode in our present and previous works using C. oleophila are
comparable with productivities that are found in literature reports,
favouring the use of the developed continuous processes for the industrial
production of citric acid in future. In view of economical aspects,
the continuous production of citric acid using yeasts shows many advantages,
compared with the traditional industrial discontinuous processes of
the last 100 years, employing selected mutant strains (several generations
of classical mutagenesis) of Aspergillus niger. The higher productivities
of presented continuous processes and the very high citric acid concentrations
that have been reached appear suitable and commercially valid for industrial
purposes, which can stand in today’s strong competition in citric acid
industry, requesting smaller fermenters in opposite to very large fermenters
of 500-
From economic point of view, integral Rj and mp are two of the most important specific fermentation parameters, determining the goodness of a fermentation process and a microbial system. A higher final integral Rj of 1.213 g/(l x hr) was reached at 80% air saturation compared with only 0.968 g/(l x hr) at 20% that corresponds to a productivity increase of about 20%. A final integral specific citric acid productivity of 0.0354 g/(g x hr) determined at 80% saturation is only at about 5.1% higher, compared with 20% saturation that reached 0.0336 g/(g x hr). It also reflects the higher biomass concentration of around 34 g/l that has been measured at 80%, compared with about 28.8 g/l (it means a stronger nitrogen limitation as well), which was determined at 20% and the compensating effect between biomass concentration and mp, determining the formation rate of the generic product Rj. Compared with the continuous fermentation process, where growth and production are running simultaneously, influencing each other (Rj results from a compensation effect between mp and biomass concentration), in batch operations they are running independent from each other. Summarizing present and previous findings, important events found in yeast process would include nitrogen depletion in fermentation medium, intracellular decrease of nitrogen content (~4%), elevation of NH4+ concentration in the cell (~37.4 mM), enhanced energy charge in cells displaying a maximum at optimum fermentation conditions (e.g. pH, air saturation and temperature), induction of specific transport system for citrate secretion, citrate secretion by active transport system, fine tuning regulation of citrate secretion by ATP, CO2, residence time etc. Furthermore, medium optimization (e.g. iron limitation etc.) increased citric acid production enormously. In view of economical aspects, continuous chemostat and repeated batch production of citric acid by yeasts seems to have many advantages compared with the traditional discontinuous industrial processes of the last 100 years utilizing Aspergillus niger. The presented results on citric acid production from glucose by yeast are the best reported in the international bibliography. I thank Professor Dr. U. Stottmeister, Mrs. E. Weissbrodt (UFZ Ctr. Envtl. Res. Leipzig-Halle, Germany) and Prof. Dr. Christian Wandrey (Institute of Biotechnology 2 of Research Center Jülich, RCJ, Germany) for their helpful advices and support. The experiments of the present manuscript comply with the currant laws of the country Germany (Institute of Biotechnology 2 of Research Centre Jülich 2 (RCJ); formerly known as Nuclear Research Centre Jülich (KFA), where the experiments were performed.
Note: Electronic Journal of Biotechnology is not responsible if on-line references cited on manuscript © 2006 by Pontificia Universidad Católica de Valparaíso -- ChileThe following images related to this document are available:Photo images[ej06004f4.jpg] [ej06004f2.jpg] [ej06004f1.jpg] [ej06004f3.jpg] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}