 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Electronic Journal of Biotechnology, Vol. 9, No. 2, April 15, 2006, pg. 107-116 RESEARCH ARTICLE Biotransformation of anthracene and fluoranthene by Absidia fusca Linnemann Danièle Villemain1, Pascale Guiraud*2, Ouahiba Bordjiba3, Régine Steiman4 1Laboratoire
Radiopharmaceutiques Biocliniques,
INSERM 0340, UFR
de Médecine de Grenoble,
Université Joseph
Fourier,
38706
Received June 6, 2005 / Accepted October 28, 2005 Code Number: ej06013 Abstract A strain of Absidia fusca was isolated from a pesticide-contaminated soil (Annaba, Algeria). The biotransformation capability of this strain towards two polycyclic aromatic hydrocarbons (PAHs): anthracene and fluoranthene was compared to that exhibited by another strain of A. fusca isolated from a non-contaminated milieu and considered as a control. The results obtained were statistically analyzed and showed that the strain isolated from the contaminated soil was more efficient than the control to remove anthracene from the medium, during all the kinetics (90% removed versus 45% after 24 hrs). Concerning fluoranthene, the amount removed by both strains was very high during the first 24 hrs however the control strain was slightly more efficient (94% versus 89%) while the results were similar for the two strains during the rest of the kinetics. This study reveals for the first time the potential interest of the species A. fusca for the bioremediation of PAHs. Keywords: Absidia fusca, anthracene, biodegradation, fluoranthene, kinetics.
Polycyclic aromatic hydrocarbons (PAHs) are fused ring aromatic compounds formed during the incomplete combustion of almost any organic material and are ubiquitously distributed in the environment (Menzie et al. 1992; Cerniglia, 1993; Chaudhry, 1994). Some of them are considered as dangerous substances because of their toxic and mutagenic or carcinogenic potentialities (Menzie et al. 1992; Nadon et al. 1995), and 16 are present on the list of priority pollutants established by the US Environmental Protection Agency (EPA). The presence of PAHs in contaminated soils and sediments poses a significant risk to the environment and human health. PAHs are hydrophobic compounds, whose persistence within ecosystems is due chiefly to their low aqueous solubility (Chaudhry, 1994). Fluoranthene is the most abundant PAH in the environment, and is so considered as a pollution indicator (Chaudhry, 1994). Anthracene is another model compound for PAHs degradation studies: its structure is found in carcinogenic PAHs such as benzo (a) pyrene and benzo (a) anthracene (Müncnerova and Augustin, 1994). Microbial biotransformation is a major environmental process affecting the fate of PAHs in both terrestrial and aquatic ecosystem (Mahmood and Rama Rao, 1993; Kästner and Mahro, 1996). A large number of bacteria that metabolize PAHs have been isolated (Alcaligenes denitrificans, Rhodococcus sp, Pseudomonas sp, Mycobacterium sp.) (Cerniglia, 1993; Harayama, 1997; Dean-Ross et al. 2001; Moody et al. 2001). A variety of bacteria can degrade certain PAHs completely to CO2 and metabolic intermediates (Kelley et al. 1993; Müncnerova and Augustin, 1994). Knowledge on fungal degradation is more limited: PAHs are oxidized to phenolic metabolites by co-metabolic process (Cerniglia, 1993; Paszczynski and Crawford, 1995; Cerniglia, 1997; Harayama, 1997). Non specific oxidation reaction catalyzed, by extracellular enzymes of white rot fungi, lead to the formation of a variety of quinones and hydroxylated aromatic compounds (Hammel, 1995; Bogan and Lamar, 1996;Tekere et al. 2005). A detailed investigation on fungal metabolism of fluoranthene was realized in Cunnighamella elegans (Müncnerova and Augustin, 1994). Preliminary works have been done in our team to evaluate the degradation or transformation of anthracene, fluoranthene, and pyrene by selected soil fungi (Krivobok et al. 1998; Salicis et al. 1999; Ravelet et al. 2000). Further studies have been recently conducted to evaluate the potential induction of the biodegradation capabilities towards anthracene and fluoranthene in fungi isolated from a contaminatedmilieu (Giraud et al. 2001). In addition, we have recently shown that anthracene can be toxic to some soil fungi (Bonnet et al. 2005). The genus Absidia is not known as efficient in the bioremediation of environmental pollutants. To our knowledge, there has been no report on the metabolism of xenobiotics by Absidia fusca Linnemann, except our recent reports about herbicides (Bordjiba et al. 2001) and phenolic compounds (Guiraud et al. 2003). In this study, the purpose was to investigate the ability of Absidia fusca to degrade these two compounds. Two strains were compared: one from the CBS-Baarn collection (normal forest soil), the other isolated from a polluted soil, and the results of the degradation kinetics obtained were statistically analyzed. Anthracene (AC) and fluoranthene (FA) were purchased from Sigma-Aldrich (Sigma Chemical Co., St Louis, MO, USA). Agar and malt extract were respectively from Coopérative Pharmaceutique Française (Melun, France) and Difal (Villefranche-sur-Saône, France). Other products were from Prolabo (Paris, France). The solid
medium used for maintaining the cultures contained: malt extract Absidia
fusca Linemann (A) was isolated in our laboratory from a polluted soil
from Algeria (Bordjiba et al. 2001) while A. fusca Linnemann
(B) (CBS 102.35, Baarn, Hollande) was from soil from pine forest (Germany).
The contaminated soil from Algeria came from a parcel treated by herbicides
and fungicides (among which the herbicides: metribuzin, metobromuron, linuron,
and metamitron, associated to the fungicides: propineb, maneozeb, and sandofan)
where tomato and potato were cultivated (El Krous, To obtain
sufficient inoculum for liquid medium cultures, the strains were grown for
1-2 weeks on MEA medium at Liquid
media with mycelia, containing AC or FA were extracted with one volume of
bidistilled ethyl acetate (25 ml) by rotary shaking at 250 rpm for 5 min
and then at 180 rpm for 30 min. Mycelia were filtered off and rinsed with
ethyl acetate. The extraction was repeated twice. The organic phases were
pooled, dried over anhydrous Na2SO4 and evaporated
to dryness at Evaluation of the level of xenobiotic transformed Residual amounts of AC and FA were calculated by the integrator connected to the HPLC system, with reference to a standard. The overall biotransformation efficiency (%) was obtained after correction with regard to the abiotic degradation (not exceeding 5%) and the extraction yield (ranging from 95 to 98%). These values were then converted into mg of xenobiotic transformed per g of dry mycelium. The treated data correspond to the amount of xenobiotic transformed expressed as mg/g of dry mycelium. Reported results are means (M), standard errors of the means (SEM) andcoefficient of variation (CV%). For each
xenobiotic a multifactorial analysis of variance was run, the factors being:
strain S (two levels: A and B), temperature T (two levels: Four groups
were obtained when crossing the factors S and T: A, In all analyses the significance level α was 0.05 and p values were considered significant at values below or equal to 0.05. Kinetics of anthracene biotransformation by A. fusca The kinetics
of AC transformation were run during five days for the four groups: A, The chromatographic profiles revealed three peaks for both strains with retention times ranging from 0.5 and 2 min while the peak corresponding to AC was observed at 5 min in our experimental conditions (data not shown). The height and the area of the peaks increased during the first 24 hrs and were higher for the strain A, while the peak corresponding to AC decreased at the same time. These peaks were not observed in the HPLC profiles of control extracts corresponding to the medium added by AC without fungus and the medium with fungus but without AC. All these observations strongly suggested that these three peaks corresponded to metabolites derived from AC. Moreover these peaks decreased after 24 hrs suggesting a biodegradation process. The percentages
of AC transformation were very high after 24 hrs with the strain A: mean
values were 89% (CV = 10.6%) at The coefficients of variation were high at 24 hrs for all groups, and decreased after 24 hrs. They were mostly high for strain B during the first 72 hrs, reaching 40% at 24 hrs (data not shown). Results
were then analyzed in order to determine the influence of the factors S and
T on AC biotransformation. Table 1 gives the results
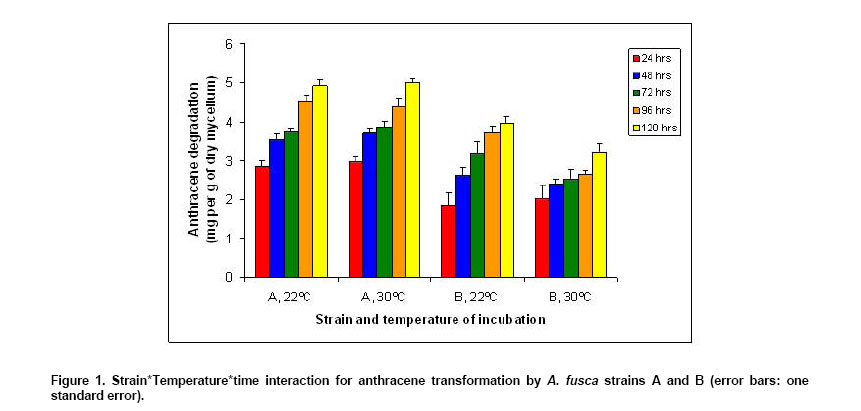
of the multifactorial variance analysis. Figure 1 illustrates
the S*T*t interaction (not significant, p = 0.35) and shows that strain A
was more efficient than strain B for AC transformation, whatever the time
and the temperature considered, this was confirmed by the demonstration of
a significant strain effect (p < 0,0001). The factor T had no effect on
strain A, whatever the value of the factor t, although the temperature effect
was significant (p = 0.012). The effect of the temperature depended on the
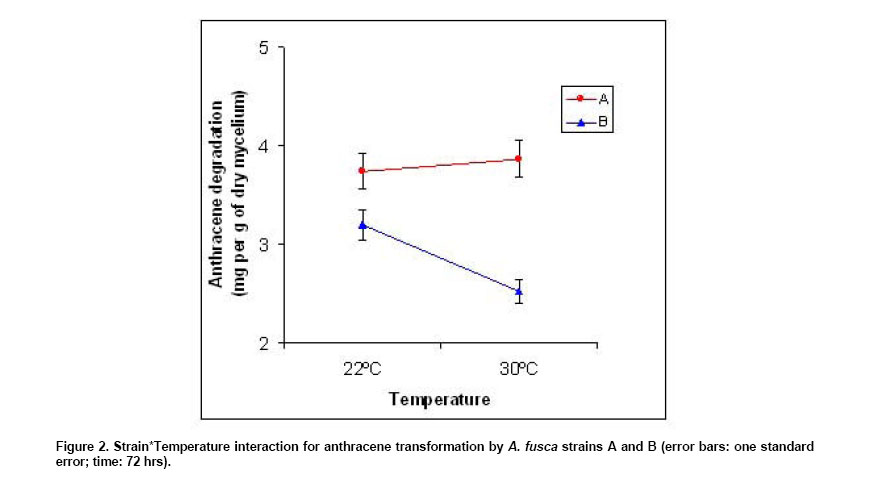
strain since the S*T interaction was significant (p = 0.001). This interaction
reported in Figure 2, showed that the mean amount of
AC degraded by strain A was almost identical at
Influence of the time of incubation on anthracene biotransformation The results
of this multifactorial analysis showed the influence of the factors S and
T on AC transformation. The results were separately studied for the four
groups: A, An analysis
of the regression variance showed for each group, a positive and significant
regression of the amount of AC transformed according to the time of incubation
(p < 0.0001 for the groups A, The four
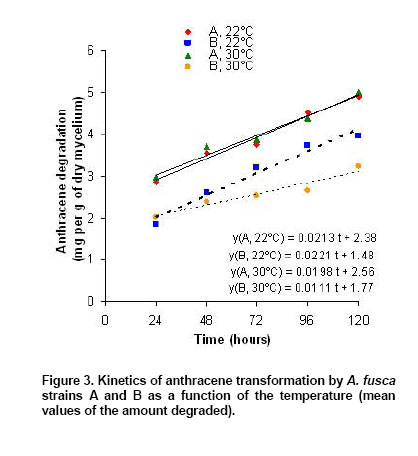
kinetics are represented in Figure 3. The slopes (rate
of AC transformation) were 0.0213, 0.0221, 0.0198 and 0.0111 mg/g of dry
mycelium/h respectively for the groups A, For the
three first groups, these slopes were not statistically different (p ≥ 0.54),
and were around 0.50 mg/g of dry mycelium/d. For strain A from the contaminated
sol, the kinetics run at Kinetics of fluoranthene biotransformation by A. fusca The kinetics of FA transformation were run as for AC. Eight replicates were done for each time. The chromatographic profiles revealed four major peaks for both strains with retention times ranging from 0.9 and 2.1 min and two smaller peaks at 3.2 and 4 min while the peak corresponding to FA was observed at 11 min in our experimental conditions (data not shown). The height and the surface of the peaks increased during the first 24 hrs, while the peak corresponding to FA decreased at the same time. These peaks were not observed in the HPLC profiles of control extracts corresponding to the medium added by FA without fungus and the medium with fungus but without FA. All these observations strongly suggested that these peaks corresponded to metabolites derived from FA. Moreover these peaks decreased after 24 hrs suggesting a biodegradation process. The percentages
of FA transformation were very high after 24 hrs: mean values were 89.3%
(CV = 4.3%) for the group A, The coefficients
of variation were all below 10% (min = 4.8%, max = 9.2%). They were not very
different according to the time in each group and also very similar from
one group to another (mean values: 6.7% for A, Influence of the strain and the temperature on fluoranthene biotransformation The results
of the multifactorial analysis are given in Table 2.
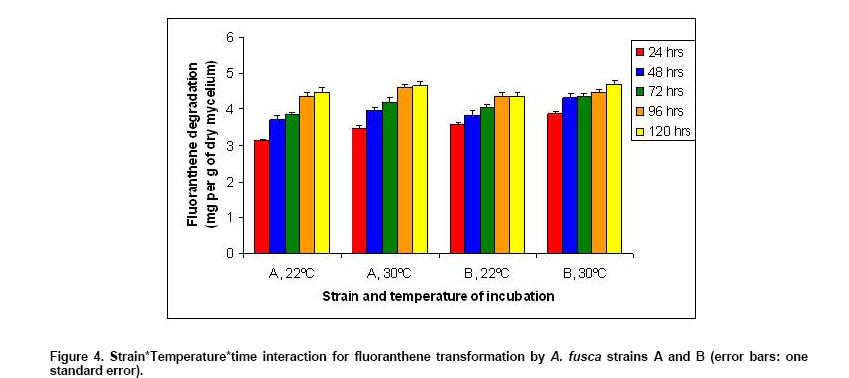
The study of the interaction S*T*t (not significant, p = 0.71) indicated
that the amount of FA transformed was always (independently of the strain
and the time considered) lower at
The strain
effect was significant (p = 0.0007), but has to be related to the significant
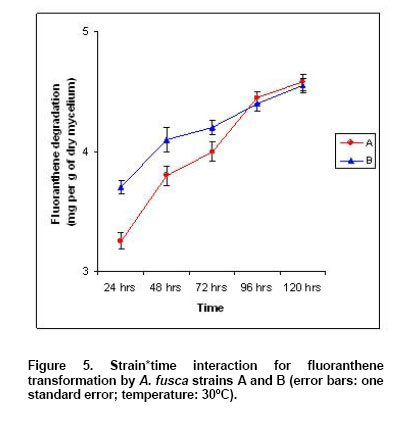
S*t interaction (p = 0.002) shown in Figure 5. The
graphic indicates that the mean amount of FA transformed was lower for strain
A during the first 72 hrs (3.29 mg/g of dry mycelium versus 3.72 at 24 hrs,
3.83 versus 4.06 at 48 hrs, 4.01 versus 4.19 at 72 hrs). The differences
between the two strains decreased during this time and then an inversion
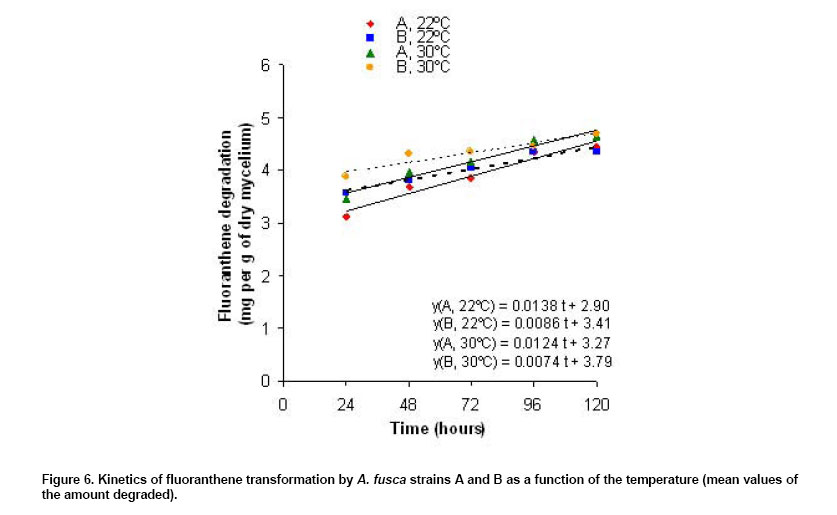
was observed (4.46 versus 4.40 at 96 hrs, 4.55 versus 4.52 at 120 hrs). For Influence of the time of incubation on fluoranthene biotransformation An analysis of the regression variance showed a positive and significant regression of the amount of FA transformed according to the time of incubation (p < 0.0001 for the four groups). The four
kinetics are represented in Figure 6. The slopes were
0.0138, 0.0123, 0.0086 and 0.0074 mg/g of dry mycelium/h respectively for
the groups A, For strain
A, the kinetics were parallel at However the kinetics of FA transformation was significantly faster for strain A than for strain B, the rate was increased by 1.6 time (difference between the slopes = 0.005 mg/g of dry mycelium/h, p = 0.01). These results were in agreement with those obtained with the multifactorial variance analysis and with the “a posteriori” tests. Comparison of the kinetics of anthracene and fluoranthene biotransformation For strain
A, the kinetics were faster for AC than for FA both at The present study reveals the potential of the species A. fusca for the transformation / degradation of two PAHs: AC and FA, and the modification of its efficiency according to the habitat it was isolated from. For both AC and FA bioremediation assays conducted with A. fusca: the differences in efficiency observed between the strains, the occurrence of new peaks on the HPLC profiles after 24 hrs of incubation, the progressive decrease of these peaks between 24 hrs and 120 hrs, together with the decrease of the peak corresponding to the xenobiotic, were an indication of the biodegradation activity of A. fusca. Identification of the metabolites produced was not undertaken in this work. Concerning
AC, the strain of A. fusca from the contaminated soil (strain A) was
considerably more efficient than strain B, since the mean amount of AC transformed
was two times higher. The incubation at Concerning FA, the mean amount transformed was slightly but significantly lower for strain A during the first 48 hrs. The transformation rate exhibited by strain A was higher than that observed for strain B. No differences in the transformation rates were obtained as a function of the temperature both for strain A and B. On the whole, the transformation rate was higher for AC than for FA with both strains (1.5 to 2.6 times). The high levels of transformation observed after 24 hrs was not a specific feature since it has been reported in the literature for numerous fungi and xenobiotics. Previous work by Giraud et al. (2001) showed that FA was more easily degraded by fungi than AC, in this work this was the case for strain B coming from a collection. Other studies showed similar results with the two molecules (Krivobok et al. 1998; Salicis et al. 1999) and this was here observed for strain A isolated from a contaminated soil. Most reports pointed out the high efficiency of the Zygomycete group for PAHs degradation, mostly Rhizopus arrhizus and the Cunninghamella genus (Cerniglia, 1993; Krivobok et al. 1998; Salicis et al. 1999), but also Absidia cylindrospora (Giraud et al. 2001). In this work we showed that A. fusca is also an interesting species with regard to its PAHs transformation / degradation capability. Fungal biotransformation and/or degradation of environmental xenobiotics, and particularly PAHs, has been extensively studied in the Basidiomycete group and particularly the white-rot fungi (Pointing, 2001; Tekere et al. 2005). In this work we confirm that some Zygomycetes can also be very efficient. In two previous reports we showed that A. fusca can degrade efficiently a wide spectrum of xenobiotics: different classes of herbicides (mostly metribuzin and metobromuron), phenolic compounds (mostly ferulic acid), pentachlorophenol (Bordjiba et al. 2001; Guiraud et al. 2003). In both studies, the biodegradation capabilities were shown to be considerably increased in the strain isolated from a polluted milieu (strain A). Here we observed that A. fusca is also efficient in the degradation of PAHs such as AC and FA, with again an increased capability in strain A at least for AC degradation. Moreover strain A was less sensitive to the temperature changes. The polluted environment has probably favoured the selection of a strain expressing low specific but efficient enzymatic systems able to degrade a large panel of molecules. This work underlines the interest of studying the microbial populations able to adapt in polluted ecosystems for bioremediation purpose.
Note: Electronic Journal of Biotechnology is not responsible if on-line references cited on manuscripts are not available any more after the date of publication. © 2006 by Pontificia Universidad Católica de Valparaíso -- ChileThe following images related to this document are available:Photo images[ej06013f2.jpg] [ej06013f5.jpg] [ej06013f4.jpg] [ej06013f6.jpg] [ej06013f3.jpg] [ej06013f1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}