 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Electronic Journal of Biotechnology, Vol. 9, No. 2, April 15, 2006, pg. 117-126 RESEARCH ARTICLE Expression of Bacillus thuringiensis insecticidal protein gene in transgenic oil palm Mei-Phing Lee†1, Li-Huey Yeun2, Ruslan Abdullah*3 1Oil Palm
Biotechnology Group,
School of Bioscience and Biotechnology,
Faculty of Science and Technology,
Universiti Kebangsaan Malaysia,
43600 Bangi,
Selangor, Malaysia,
E-mail: phing_san@yahoo.com Financial support: Ministry of Science, Technology and Innovation, Malaysia (IRPA 01-02-02-0168 and IRPA 09-02-02-0033. Received August 1, 2005 / Accepted October 4, 2005 Code Number: ej06014 Abstract Oil palm, like all other crops, is susceptible to attack from several insect pests causing significant reduction in productivity. In the past, cry genes from Bacillus thuringiensis have been reported to be effective in conferring resistance towards insect pests in crops such as corn and rice. One of the advantages of these toxin proteins is their specificity towards certain harmful insects. A rapid and efficient method was developed for the transformation and evaluation of CryIA(b) expression in oil palm. A recombinant vector was introduced into immature embryos (IEs) of oil palm via the biolistic method. More than 700 putative transformed IEs from independent transformation events were generated. Transient transformation efficiency of 81-100 % was achieved. We found that pre-treatment of target tissues with phytohormones is essential for increasing the transformation efficiency. This finding could enable higher transformation rate in oil palm that was previously difficult to transform. PCR analysis further confirmed the presence of the CryIA(b) in the transformed tissues. Expression of CryIA(b) from PCR-positive samples was further confirmed using a rapid gene expression detection system. This novel and rapid detection system could serve as a good opportunity to analyze the impact of transgenes upon transfer to the new environment, especially for crops with long generation cycle, such as oil palm. Keywords: CryIA(b) gene, gene expression, insect resistance, oil palm transformation, rapid detection system, transgenic oil palm.
Oil palm (Elaeis guineensis Jacq.) is the source of the most sought-after edible oil in the world market. With the advent of new biotechnology tools, in particular gene manipulation, palm oil monopoly as a commodity is challenged by other oil-bearing crops such as soybean (Kinney et al. 2001), sunflower seed (Osorio et al. 1995) and rape seed (Ramachandran et al. 2000; Dehesh et al. 2001) that have been successfully modified genetically. Thus, it is imperative to increase oil palm yield to meet the ever-increasing global demands and also to expand its uses. In the past, conventional breeding of oil palm is a low-efficiency and time-consuming process due to its long reproduction cycle and slow seed maturation (Mayes et al. 2000). Current conventional breeding programmes for oil palm are supported by modern biotechnology approaches such as the use of tissue culture, genetic transformation and marker-aided selection, to further improve oil palm commercial value and productivity (Mayes et al. 2000). Over the past 10 years, our group has been actively involved in genetic engineering of oil palm (Abdullah et al. 2003; Abdullah et al. 2005). Although many studies have been done, but there are very few publications on genetic manipulation of oil palm, probably due to the high commercial implication tagged to the crop and also due to the crops own physiological characteristics such as long generation cycle. Special emphasis was placed initially in developing techniques for genes transfer into oil palm, both using direct gene transfer (DGT) and Agrobacterium-mediated approaches. Both techniques were then used to transfer useful genes such as cowpea trypsin inhibitor (CpTI) (Abdullah et al. 2003) and Bacillus thuringiensis (Bt) crystal insecticidal protein genes, to address problems related to insect pests (Sharma et al. 2000; Sharma et al. 2002) and chitinase (to address problems related to basal stem rot). Transgenic plants such as tomato, tobacco, cabbage and rice containing CryIA(b) gene have been obtained using both DGT and Agrobacterium transformation methods (Cheng et al. 1998; Jouanin et al. 1998; Robinson, 1999; Bhattacharya et al. 2002). In this article, we report successful transformation of oil palm using particle bombardment for expression of an agronomic trait. We have carried out the construction of the CryIA(b) gene cassette (Sardana et al. 1996) which have been modified to express highly in plants, particularly oil palm. In the investigation, the synthetic CryIA(b) was introduced into oil palm, data for successful transformation and expression of the gene within the oil palm tissue were also presented. The CryIA(b) gene when expressed produced proteins which upon crystallization are highly toxic to Lepidoptera (Masson et al. 1999; Reardon et al. 2004), among which Metisa plana is a major insect pest for oil palm (Wood, 1968). Using our established DGT methods for oil palm, we then developed a rapid detection system for evaluating transgene expression among putative transformed tissues. The procedure developed is very sensitive, rapid and eliminates the long waiting period for transgenic plants to reach maturity. Escherichia
coli strain DH5α was used as plasmid host for the construction
of transformation vectors. Standard cloning techniques (Sambrook
et al. 2001) were used to construct the recombinant plasmid, pRMP as
shown in Figure 1. The coding sequence for CryIA(b) gene,
a derivative of plasmid pUBB (Prof. Illimaar Altosaar, University of Ottawa,
Canada), and known to produce lepidopteran-specific δ-endotoxin, was
inserted into pCAMBIA 1301 cloning vector (Dr. Richard A. Jefferson, CAMBIA,
Australia) using the BamHI site. The HindIII site was then
deleted through the excision of the Oil palm
(E. guineensis Jacq. var. tenera) samples were obtained from
Pamol Plantations Sdn. Bhd. IEs were isolated from oil palm fruit bunch harvested
9-11 weeks after anthesis (WAA) according to protocols described earlier
(Abdullah et al. 2005). Extracted IEs were cultured on N6 media
with or without 2.5 mg/l 2,4 D and incubated either in the dark at 28 ± Pre-cultured
IEs were bombarded using the PDS-1000/He particle delivery system (BioRad) following the procedure established
elsewhere (Wright et al. 2001) with some modifications
(Abdullah et al. 2005). All components of the chamber were
surface-sterilized using 70% (v/v) ethanol prior to bombardment. Plasmid
DNA (2.5 μg) were coated onto 3 mg of resuspended gold particles. This
was followed by the addition of Isolation of genomic DNA and total RNA Genomic DNA and total RNA was isolated according to modified protocols of Doyle and Doyle (1990) and Verwoerd et al. (1989), respectively, using bombarded IEs 3 days after bombardment. Both DNA and RNA were quantified using a spectrophotometer. Expression analysis of CryIA(b) gene in IEs Oligonucleotides
were designed to amplify region specific to the CryIA(b) coding sequence.
For the purpose of evaluating transformation events, evidence of transgene
presence is considered positive PCR or a positive expression assay for the
gene of interest. All PCR and RT-PCR were carried out according to the manufacturer’s
instruction manual (Promega).
The primers set used were as follows: CRYF1 5’~GGTTCGTTCTCGGACTAGTT and CRYR1
5’~CTGGTAAGTTGGGACACTGT. A total of 0.1-0.4 μg genomic DNA was used
as template in a 50 μl PCR reaction mix containing 25 pmol of each primer,
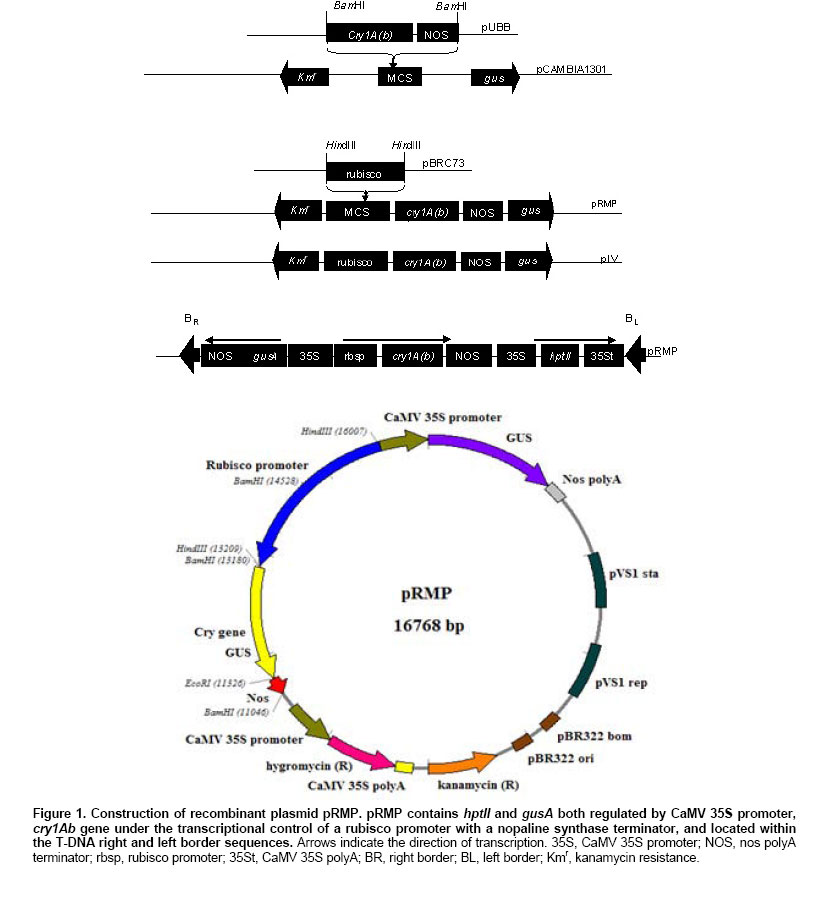
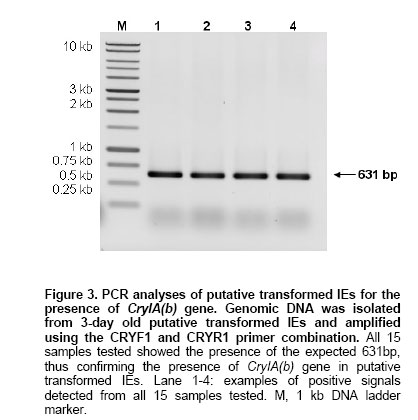
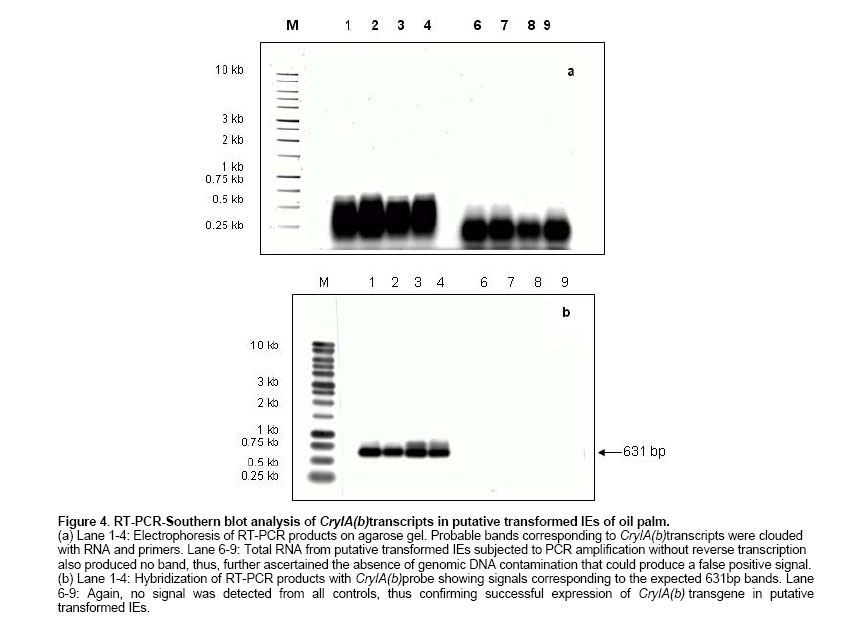
2.5 U Taq DNA polymerase, Characterization of transformation vector The open reading frame (ORF) of CryIA(b) was successfully cloned into pCAMBIA 1301 under the control of rubisco promoter in the sense orientation creating a new recombinant plasmid, pRMP (Figure 1). The orientation and integration of CryIA(b) in pRMP were confirmed through restriction mapping, Southern blotting and nucleotide sequencing. Southern blot analysis following multiple enzymatic digestions showed that CryIA(b) was successfully integrated into pRMP. Integration of CryIA(b) inpRMP was in-frame as confirmed through nucleotide sequencing with only the BamHI site connecting the rubisco promoter with the CryIA(b) coding sequence. Besides the CryIA(b) gene which is under the control of a rubisco promoter, the newly constructed pRMP also contains hptII (selectable marker gene) and gus (reporter gene), from the original pCAMBIA1301. Since all three genes cassette are flanked by the T-DNA right and left border sequences, thus, pRMP is useful for both biolistic-mediated and Agrobacterium-mediated transformation experiments where they should be transferred in the process of successful gene transfer mediated by Agrobacterium. Pre-treatment of target tissues In our earlier work, we have shown that 2,4-D pre-treatment of target tissues prior to particle bombardment and Agrobacterium infection (Abdullah et al. 2005) was essential for increasing the number of transformation events in target cells or tissues. IEs pre-cultured on N6 media supplemented with 2.5 mg/l 2,4-D prior to bombardment also showed different GUS expression pattern as compared to IEs maintained on hormone-free N6 media (Figure 2). In contrast, no blue spot or shade was observed on all control or non-bombarded tissues. The GUS-positive blue spots on IEs cultured on hormone-free N6 media were distinct, isolated and localized, whereas the GUS stains on 2,4-D pre-treated IEs spread over a larger area. There are two possibilities that may result in the expression of GUS that produced the blue stains over a larger area. First, the spread could be a consequence of a possible leakage between cells following damage caused during the process of bombardment. However, if this were the case, similar GUS expression pattern would also be observed on IEs pre-treated on hormone-free N6 media since both samples were treated the same. Second, the spread of blue stains may be from relatively more cells that may have received the gusA gene either through gene transfer (bombardment) or daughter cells derived from transformed cells undergoing cell divisions. This larger population of cells now possessing the gusA gene may have resulted in larger area of the tissues being stained blue following the expression of the transferred gene on the substrate. Therefore, it is more likely that the 2,4-D present in pre-treatment media must had influenced the expression pattern of the gusA gene transferred as observed in Figure 2. Being an auxin (Jain and Minocha, 2000; Pasternak et al. 2002), 2,4-D induces rapid cell division. Since the target tissues were pre-treated with 2,4-D prior to bombardment, therefore, upon successful transfer and integration, the gusA gene could easily be amplified together during subsequent cell divisions, thus giving rise to a wider spread of the blue stains observed (Abdullah et al. 2005). On the other hand, without 2,4-D, IEs pretreated on hormone-free N6 media did not undergo rapid cell division, but following the path of differentiation instead. Thus, the gusA stains detected in untreated IEs were isolated and distinct. In addition the stains were mainly localized in cells that may have received the transgene from bombardment only. Similar observations were also reported in rice transformation using Agrobacterium, where only target tissues cultured on 2,4-D-containing media were capable of expressing GUS whereas those untreated tissues did not (unpublished data). Yang et al. (1999) also claimed that cell divisions correlate with maximum transformation events, suggesting cell division is essential for cell competency. Competent cells have been associated to its susceptibility to receive foreign DNA during the process of transformation (Tang and Tian, 2003). Though we observed similar results in the case of oil palm, the reasons still remain unclear. However, it is believed that during cell division, active replication of DNA strand takes place. This is when chromosomes unwind and the double helix denatures (Gerald, 2002), hence exposing the fragile DNA. It is assumed that it was at this time that cells are most vulnerable to the invasion of foreign DNA. Illegitimate recombination between the vulnerable DNA and transgenes may have resulted in stable integration (Tzfira and Citovsky, 2002; Hohe and Reski, 2003). Therefore, factors that induce rapid cell division could play major role in ensuring higher transformation events. Under our new experimental design, putatively transformed IEs were assayed 3 days after transformation. 750 IEs bombarded with pRMP were randomly selected and analyzed through histochemical staining (Figure 2). Transient GUS assay was used to evaluate the efficiency of gene transfer, where transformation efficiency is defined as the number of independent event recovered per explant bombarded (Wright et al. 2001). This was determined using result from the GUS assay. GUS-positive blue spots or stains were detected on randomly selected IEs assayed, suggesting gene transfer had taken place. In contrast, no blue spot was observed in all control samples assayed. Here, observation of one or more blue spots on a single IE is considered as one unit of expression (Klein et al. 1988). On average, the transient transformation efficiency of IEs bombarded once was recorded at 81% as compared to 100% for those subjected to double bombardment. Double or multiple bombardments is not normally practiced in other crops, as it would cause excessive wounding to target tissues (Jiang et al. 2000) resulting in eventual death. However, we believed that oil palm being a hardy plant could withstand certain amount of wounding to its tissues. Post bombardment observation showed that there was no excessive accumulation of phenolic compound in the culture medium, which would be the case for severely injured or dying oil palm tissues. IEs from both treatments were cultured on hormone-free N6 media and regenerated to complete plantlets (Figure 2). Apart from about 3% losses due to contamination, all bombarded IEs were regenerated into complete plantlets. The main objective of direct plant regeneration was to evaluate the impact of double bombardment on the plant as compared to single bombardment. Although the construct contained the hptII gene, antibiotic selection was not performed, as this would cause cell death to untransformed cells, thus inflicting additional stress to the limited transformed cells present. During antibiotic selection, dying cells inhibit nutrient supply to transgenic cells or excrete toxic compounds, which further impedes proliferation of transgenic cells to differentiate into transgenic plants (Ebinuma et al. 2001). This adverse effect would then influence IEs growth thus causing difficulties in data collection. Preliminary observation showed that there were no morphological differences between plantlets derived from IEs bombarded once or twice (Figure 2e). Similar observations were also noted in the formation of shoots and roots of plants from both groups. Plants from both groups also exhibited similar growth rate. Therefore, based on these observations, oil palm IEs can withstand double bombardment and would still be able to regenerate into healthy plantlets. However, these qualitative observations need further detailed study, especially on the plants performance upon maturity. Detection of transgene using PCR Many techniques have been developed to detect transgenes in T0 plants. These include PCR-based techniques followed by gel electrophoresis and detection, and are routinely used to detect transgenes in plants (Higuchi et al. 1992; Abedinia et al. 1997). Other techniques such as the use of molecular beacon assays (Kota et al. 1999) and FISH (Jin et al. 2002) had also been used. But this was not the case for slow growing plants like oil palm where detection of transgenes has always been a major problem. In this study, preliminary analyses on transformants were carried out by PCR analyses. Two primers, CRYF1 (nt 170-189) and CRYRI (nt 800-781) designed based on the coding sequence of CryIA(b) from plasmid pUBB with an expected PCR product of 631 bp were used in subsequent PCR analyses. Genomic DNA isolated from fresh tissues of putative transformed and non-transformed (negative control) plants were used as templates in the analyses. In all cases, the expected 631 bp bands were present in all 15 samples tested (Figure 3), indicating the presence of CryIA(b) in the genomic DNA of putative transformed plants. In contrast, no band was observed for all control samples. The experiments were repeated three times, giving the same result, suggesting successful gene transfer into oil palm via particle bombardment. The 100% transient transformation efficiency observed following PCR of CryIA(b) was similar to those observed earlier from GUS assay carried out using the same sample. Therefore, this further confirmed successful transfer of transgenes from pRMP into the oil palm tissues. However, the presence of CryIA(b) in putative transformed plants is no indication of its functionality in its new environment. Expression of CryIA(b) gene in oil palm Evaluation of transgene expression in T0 transgenic plants has always been difficult, especially when the transgenes undergoes various modifications in its new environment (Stam et al. 1998; Francis and Spiker, 2005). Many reports have shown that transgenes may be present in its new environment but were not expressed or expressed at a very low level making it undetectable. Failure for transgene to express may be due to mutations in the transgenes (Tinland, 1996), post-transcriptional gene silencing (Mitsuhara et al. 2002; Szittya et al. 2003), or chromatin-related transgene silencing (Francis and Spiker, 2005). In this study, although PCR analyses could easily detect the presence of CryIA(b) in the putative transformed plants, but attempts to detect its expression and functionality could not be easily carried out. Several attempts using Northern blots to detect specific RNA sequences corresponding to the CryIA(b) gene was unsuccessful (data not shown). Subsequently, however, RT-PCR analyses produced signals but could not be discriminated between those of the transgene [CryIA(b)] or contaminating primers and RNA (Figure 4a). However, when the RT-PCR products were blotted onto nylon membrane and hybridized against the CryIA(b) probe, it gave rise to positive signals with the expected size (631 bp, Figure 4b). The RT-PCR products were subsequently sequenced and showed that they were 100% homologous to the CryIA(b) from the original plasmid pUBB (Prof. Illimaar Altosaar, University of Ottawa, Canada). Thus, the presence of CryIA(b) mRNA transcripts shows that it is fully functional in oil palm, at least at the transcriptional level. Therefore, it can be concluded that the DNA and RNA specific to CryIA(b) were actually present in putative transformed oil palm (as also shown in PCR analyses) but were below detectable level. However, once amplified (using RT-PCR), these could easily be detected as RT-PCR allows for reverse transcription of mRNA and amplification of the transcripts (Pfaffl and Hageleit, 2001). Combining Southern blot with RT-PCR further substantiate evidence on the functionality of CryIA(b) in oil palm. These observations are the first evidence on the functionality of a transgene in oil palm following transformation and could also serve as a sensitive detection assay as shown by the positive signals produced. Most published papers on oil palm transformation (Chowdhury, 1997) used gusA, bar or ppt as reporter genes, which are easy to assay. In addition, there has been no conclusive report on transgenes expression in oil palm to date. In this regard, the fact that the CryIA(b) transcripts were detected directly using the RT-PCR-Southern Blotting method, it shows that not only the gene has been successfully transferred but is also successfully transcribed in its new environment. The combination of RT-PCR-Southern Blotting method is therefore useful to detect and study the fate of transgenes upon transfer in its new environment. This is essentially more important when multi-cellular tissues were used as target tissues, where only a minute fraction of cells within the tissues would be transiently transformed per bombarded explant. In such cases, it is quite impossible to determine the fate of the newly transferred transgene in its new environment. Similarly, it is often difficult to quantify transgene copy number in its new environment (Mason et al. 2002; Bubner et al. 2004). However, this would not be the case for transgenic plants that are derived from single cells, where every cell in the plant would have the stably integrated transgene. Similar findings have also been observed in maize transformed with Bt gene, where the expression of reporter genes at the protein level is indirectly implied. This led to the development of a direct detection method for protein expression of the gene of interest (Sardana et al. 1996). However, our approach gives a more accurate picture on the of transcription level of the gene of interest in oil palm. For example, it was found that, the CryIA(b) transcripts were relatively little compared to the GUS protein expression observed in the transformed IEs. This approach could also serve as an important tool in promoter study, as it would be more precise to evaluate promoter efficiency at the transcription level as compared to the protein level, since it will not be influence by translational factors. In conclusion, this paper provides the first evidence on the functionality of a transgene in oil palm following transformation. In addition, this detection technique that combines RT-PCR and Southern blotting is highly sensitive in detecting minute traces of transgenes expression in putative transformed plants. It is a useful a tool that enable closer expression studies of individual genes at the cellular level where transcripts analyses could be carried out immediately following transformation. This is especially important for crops with slow growth rate such as oil palm. In addition, the system also allows for the detection of mRNA expression reflecting biological phenomena in minute amount of cells and also for accurate evaluation of essential but lowly expressed mRNA in transgenic plants. The authors would like to thank Prof. Illimaar Altoosaar, Ottawa University, Canada, for the cry1A(b) gene; Dr. Richard A. Jefferson, CAMBIA, Australia, for the pCAMBIA1301 cloning vector; Dr. J. K. Kim, Myongji University, Korea, for the rubisco promoter and Universiti Kebangsaan Malaysia (UKM) for research facilities.
Note: Electronic Journal of Biotechnology is not responsible if on-line references cited on manuscripts are not available any more after the date of publication. © 2006 by Pontificia Universidad Católica de Valparaíso -- ChileThe following images related to this document are available:Photo images[ej06014f4.jpg] [ej06014f2.jpg] [ej06014f3.jpg] [ej06014f1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}