 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Electronic Journal of Biotechnology, Vol. 9, No. 2, April 15, 2006, pg. 133-143 RESEARCH ARTICLE Production of polyclonal antisera to manganese superoxide dismutase expressed in downy mildew resistant pearl millet and its application for immunodiagnosis Mysore Pandurangaraj Urs Babitha1, Devaiah Madhu2, Harishchandra Sripathi Prakash*3, Hunthrike Shekar Shetty4 1,Downy Mildew
Research Laboratory,
Department of Studies in Applied Botany,
Seed Pathology and Biotechnology,
University of Mysore, Manasagangotri,
Mysore 570 006, India,
Tel/Fax: 91 821 2414450,
E-mail: dr.babitha@gmail.com Financial support: Indian Council of Agricultural Research, New Delhi and Council for Scientific and Industrial Research, New Delhi, India. Received January 31, 2005 / Accepted September 6, 2005 Code Number: ej06016 Abstract Level of superoxide dismutase (SOD) in the seedlings of downy mildew resistant pearl millet genotype (IP 18293) increased 3-fold upon inoculation with Sclerospora graminicola. Mn-SOD was purified from pearl millet by ion-exchange and gel filtration chromatography. Polyclonal antibody was raised in rabbit against Mn-SOD of pearl millet. Anti-Mn SOD had an antibody titer of 1:20,000. ELISA results revealed a 1.5-fold increase in pure Mn-SOD preparation compared to the crude preparation. The antibody reactivity as detected by ELISA revealed that Mn-SOD is higher at 4 hrs post-inoculation in resistant pearl millet seedlings. The reactivity also showed the Mn-SOD is more in roots of inoculated resistant seedlings. SOD reactivity was also determined by DIBA (Dot-immunobinding assay). The results suggested that the Mn-SOD antibody reacted more strongly with crude SOD of the inoculated resistant seedlings than inoculated resistant seedlings. Western blot analysis revealed the induction of 35 kDa SOD protein in the resistant genotype. Its appearance on western blot coincided with high SOD activity. Immunolocalization experiments showed that SOD protein was abundant in the vascular bundles. The antibody produced to Mn-SOD was specific as judged by ELISA, DIBA, western blot and immunofluorescence assays. The work described here investigates the possibility of using the serological techniques to assess the reactivity of antibody with the SOD antigen. Keywords: DIBA, downy mildew, ELISA, immunofluorescence, manganese superoxide dismutase, pearl millet, Sclerospora graminicola, Western blot.
Downy mildew of pearl millet (Pennisetum glaucum L. (R.) Br.) incited by S. graminicola (Sacc.) Schroet. is a devastating disease. The disease is systemic and the pathogen is an obligate biotroph. Its economic significance has become more accentuated than ever before, especially in relation to the high yielding genotypes (Shetty, 1990). Considerable progress has been made in understanding the interaction between the downy mildew pathogen and pearl millet. Particularly striking has been the information gained at the histological and biochemical levels (Shetty et al. 1998). Involvement of the enzymes such as lipoxygenase, phenylalanine ammonia lyase, peroxidase, ATPase, b-1,3-glucanase and RNAase in S. graminicola and pearl millet interaction has been reported (Nagarathna et al. 1992; Nagarathna et al. 1993; Sreedhara et al. 1995; Ramachandra et al. 2000; Shivakumar et al. 2000; Madhu et al. 2001). The role of the enzyme superoxide dismutase in imparting resistance to downy mildew in pearl millet has been established by the authors (Babitha et al. 2002a). Reactive O2 species (ROS) are produced in both unstressed and stressed cells. Plants have well-developed defence systems against ROS, involving both limiting the formation of ROS as well as instituting its removal (Alscher et al. 2002). Plants usually keep the levels of ROS under tight control by the production of scavenging enzymes and non-enzymatic antioxidants (Wojtaszek, 1997; Kuzniak and Urbanek, 2000; Moller, 2001; Vranova et al. 2002). Within a cell, the superoxide dismutases (SODs) constitute the first line of defence against ROS (Alscher et al. 2002). Superoxide dismutases (SODs; EC 1.15.1.1) are a family of metalloenzymes that catalyze the disproportionation of superoxide (O2- ) radicals, and they play an important role in protecting cells against the toxic effects of superoxide radicals produced in different cellular loci (Fridovich, 1995; Halliwell and Gutteridge, 2000). It has been extensively reported in the literature that the response of SOD activity and other antioxidative enzymes to oxidative stress varies according to the environmental conditions, plant tissue, stage of development, etc. (Dat et al. 2000; Alscher et al. 2002). Recent work on Cd-induced oxidative stress has confirmed such a statement (McCarthy et al. 2001; Sandalio et al. 2001; Romero-Puertas et al. 2002; Schutzendubel et al. 2002). Three classes of SODs, differing in the metals at their catalytic active site, are known in plants. The CuZnSODs are localized in the cytosol, chloroplasts, nucleus, and apoplast; the Mn-SODs in the mitochondria and peroxisomes; and the FeSODs in the chloroplasts (Bowler et al. 1994; Ogawa et al. 1996). A novel type of SOD containing nickel as a cofactor has been found in several Streptomyces spp. (Youn et al. 1996). Both O2- and H2O2 have been shown to act directly or indirectly in plant defence and signal transduction (Bolwell, 1996; Kuzniak and Urbanek, 2000; Yoshioka et al. 2001; Vranova et al. 2002). The conversion of O2- and H2O2 to OH, catalyzed by transition metals (Haber-Weiss reaction), accounts for the severe toxicity of ROS in plants (Wojtaszek, 1997). Activated
oxygen or oxygen-free radical mediated damage to plants has been implicated
in many plant stress situations. The enhanced SOD activity might increase
oxidative stress due to increased production of H2O2 (Tenhaken
et al. 1995). Increased activity of the enzyme has been reported to induce
cell dysfunction and death. Moreover H2O2 is implicated
in hypersensitive cell death, thus limiting the spread of cell death by induction
of cell protectant genes in surrounding cells. H2O2 also
inhibits the growth and viability of diverse microbial pathogens (Wu
et al. 1995). The oxidative potential of H2O2 also
contributes to plant cell wall strengthening during plant-pathogen interactions
through peroxidase-mediated cross-linking of proline-rich structural proteins
and phytoalexin biosynthesis during oxidative burst. Manganese-containing
superoxide dismutases have been characterized from a wide range of organisms
including bacteria, algae, fungi, and animals (Bannister et
al. 1987), and also from several higher plants (Streller
et al. 1994; Kroniger et al. 1995). In eukaryotic cells,
Mn-SODs have been found to be localized mainly in mitochondria from different
organisms (Fridovich, 1995; Halliwell and
Gutteridge, 2000). Mn-SODs are chiefly present in mitochondria (Halliwell
and Gutteridge, 2000), but also occur in different types of peroxisomes
(Corpas et al. 1998; del Rio et al. 1998; Differential induction of superoxide dismutase (SOD) in downy mildew-resistant and susceptible genotypes of pearl millet (Pennisetum glaucum) was observed on inoculation with Sclerospora graminicola (Babitha et al. 2002a). SOD activity was studied in resistant (IP18292) and susceptible (23B) pearl millet seedlings inoculated with S. graminicola. SOD activity increased by 2 x 3-fold in resistant seedlings upon inoculation. Native PAGE analysis showed four isozymes of SOD, three of which (SOD-1, -2 and -4) were Cu/Zn-SOD, whereas isozyme SOD-3 was Mn-SOD whose intensity increased in the resistant genotype upon inoculation. Mn-SOD involved in defence mechanism was purified from downy mildew resistant pearl millet genotype (Babitha et al. 2002b). The present investigation was aimed at production of polyclonal antibody against Mn-SOD. The antibody was used to examine the Mn-SOD protein in downy mildew defence through ELISA, DIBA, Western blot and immunofluorescence. Comparison of Mn-SOD activity with Mn-SOD protein amount which was determined immunologically indicates that the activity of the enzyme is increased in the downy mildew resistant pearl millet after inoculation with the pathogen S. graminicola. This is the first report for quantifying Mn-SOD in pearl millet with special reference to downy mildew defence. Seeds of pearl millet genotypes, resistant (IP 18293) and susceptible (HB3) to downy mildew, were obtained from ICRISAT, Hyderabad and All India Co-ordinated Pearl Millet Improvement Project (AICPMIP), Jodhpur, India. Pathogen
and inoculation of the seedlings. Seeds were germinated on moist blotter
paper discs in petri dishes at 25 ± The crude
enzyme source was prepared by homogenizing the seedlings with The enzyme Mn-SOD was purified from inoculated resistant pearl millet seedlings by homogenization, ammonium sulphate precipitation, anion exchange chromatography and gel-filtration chromatography followed by SDS-PAGE (Babitha et al. 2002b). This was used as antigen for immunization and to test the serological relationships of polyclonal antibody with pure Mn-SOD and crude SOD extract of different pearl millet genotypes with varying degrees of resistance to downy mildew disease. Antiserum
was raised against the purified Mn-SOD from pearl millet in New Zealand white
rabbit following the standard immunization protocol (Harlow
and Lane, 1988). For immunization 800 µg of protein was used. The pure
protein in phosphate buffered saline (PBS; The activity
of SOD was assayed by measuring its ability to inhibit the photochemical
reduction of nitroblue tetrazolium (NBT) using the method of Beauchamp
and Fridovich (1971). The 300 µl reaction mixture contained Enzyme linked immunosorbent assay (ELISA) Determination
of antibody titre of the Polyclonal antibody-MnSOD (PAb-Mn SOD). Indirect
ELISA was used to detect the titer of polyclonal antibody and to test their
specificity for SOD of pearl millet. ELISA was carried out in commercially
available 96-well microtiter plates (Nunc, Denmark). Antigen
(10-50 µl) was loaded into each well of the ELISA plate and the volume
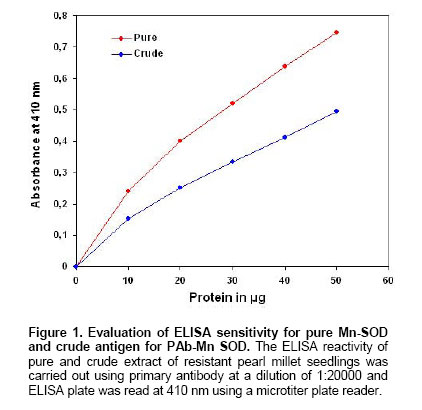
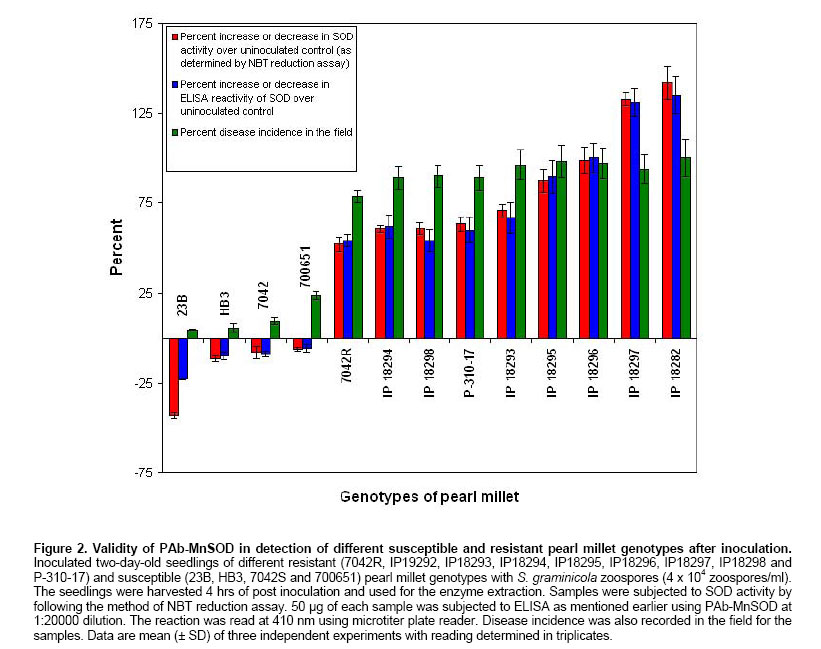
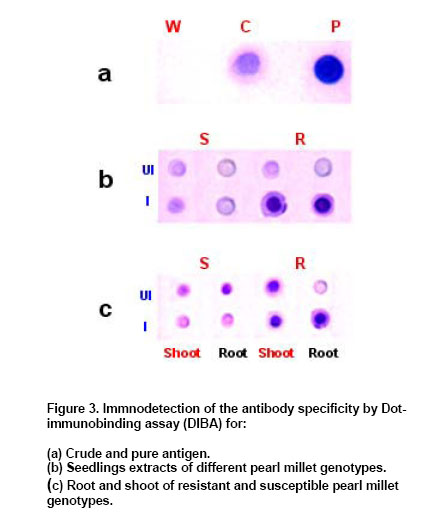
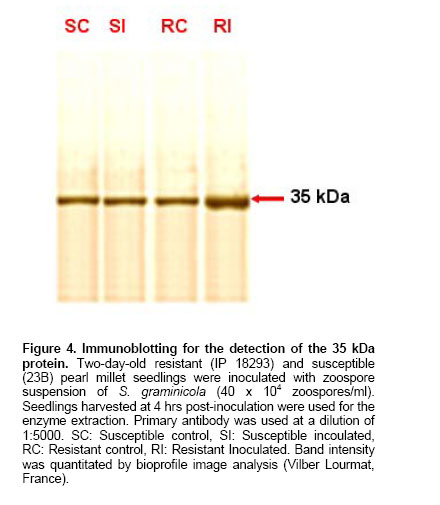
was made upto 100 µl / well with antigen buffer (PBS with Evaluation of ELISA sensitivity for pure and crude antigen for PAb-Mn SOD Dose-dependent reactivity of the PAb-Mn SOD was evaluated for crude and pure enzyme preparation at different protein concentration using ELISA test. Determination of PAb-MnSOD in resistant and susceptible pearl millet genotypes Two-day-old seedlings of resistant (IP 18293) and susceptible (23B) pearl millet genotypes were inoculated with S. graminicola zoospores. Seedlings were harvested at two-hour intervals up to 24 hrs and the enzyme was extracted. The enzyme (50 µg) was quantified by ELISA using 1:20,000 of the PAb-Mn SOD. Determination of PAb-Mn SOD in different tissues of pearl millet seedlings Root and shoot portions of seedlings were excised 4 hrs post-inoculation and the enzyme was extracted. The extract was subjected to ELISA. Validity of PAb-Mn SOD in detection of susceptible and resistant pearl millet genotypes after inoculation Two-day-old seedlings of different resistant (7042R, IP19292, IP18293, IP18294, IP18295, IP18296, IP18297, IP18298 and P-310-17) and susceptible (23B, HB3, 7042S, 700651 and ‘Kalukombu’) pearl millet genotypes were inoculated with S. graminicola zoospores. Four hours after inoculation the enzyme extract was obtained and subjected to ELISA. Dot-immunobinding assay (DIBA) for Mn-SOD Aliquots of 5 µl (3 µg) of the sample were spotted onto the nitrocellulose membrane using a dot-blot manifold apparatus. All subsequent incubations and washing steps were carried out at room temperature on a shaker at 80 oscillations per min. The membrane was placed in a glass Petri dish containing 50 to 100 ml of blocking solution (0.02% tween-20 and 5% fat free milk powder in TBS) for 30 min at room temperature. The blots were rinsed thrice in TBS containing 0.3% tween. The membrane was then incubated for 2 hr in primary antibody (1:5000) at room temperature. The blot was washed as mentioned above and incubated in secondary antibody- Alkaline phosphatase with dilution of 1:2000 in TBS and incubated for 2 hrs. The blot was then washed in TBS thrice. Finally the membrane was incubated in BCIP/NBT in dark for 10-30 min for development of colour. The blot was rinsed and air-dried. Seedling extracts of different pearl millet genotypes were tested against PAb-Mn SOD using DIBA. Two-day-old seedlings of resistant (IP 18293) and susceptible (23B) pearl millet genotypes were inoculated with S. graminicola and samples were collected 4 hrs post-inoculation for enzyme extraction. Distilled water treated seedlings served as control. 3 µg protein of each sample was used in the immunobinding assay. PAb-Mn SOD was used at a dilution of 1:5000. Tissue-specific expression of PAb-SOD in resistant and susceptible pearl millet genotypes were analyzed by immuno-detection. Root and shoot portion of two-day-old S. graminicola inoculated seedlings of resistant and susceptible pearl millet genotypes were excised and used for enzyme extraction. These samples (3 µg) were subjected to immunobinding assay. PAb-Mn SOD was used at a dilution of 1:5000. Immunoblotting for the detection of 35 kDa protein Crude
extracts of pearl millet seedlings resistant and susceptible to downy mildew
were subjected to SDS-PAGE on 12% separating and 5% stacking slab gel according
to the method of Laemmli (1970). One half of the gel was
stained with coomassie blue and other half of the gel was electro-transferred
onto a Nitrocellulose membrane (Towbin et al. 1979). After
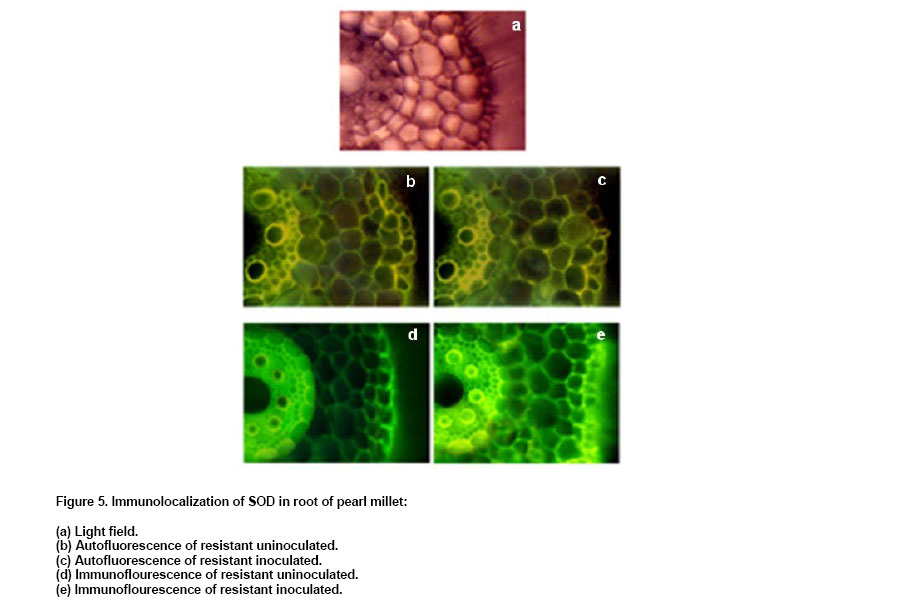
electrophoresis, the gel was allowed to equilibrate in transfer buffer ( Immunolocalization of SOD in pearl millet Two-day-old resistant (IP 18293) and susceptible (23B) pearl millet genotypes were inoculated with S. graminicola zoospores (4 x 104 zoospores/ml). Four hours after inoculation, root and shoot portions of seedlings were excised, sections were taken using a cryostat and the sections were incubated for 15 min with PBS containing 1% non-specific antibody. The sections were incubated in 1:5000 dilution of PAb-Mn SOD for 30 min, and rinsed three times in PBS (5 min). Sections were incubated in a secondary FITC conjugated anti rabbit Ig at 1:500 dilution in PBS for 30 min and rinsed three times in PBS (5 min.). The antigen-antibody reaction in the tissue was observed under Wild Leitz fluorescence microscope attached with photoautomat. Appropriate controls (absence of secondary antibody) were setup for each experiment. Polyclonal antibody against Mn-SOD was obtained after intramuscular injections. The antibody sensitivity and the titre were determined using ELISA. The antibody titre was found to be 1:20000. The antibody reactivity was 1.5-fold more in pure sample than that of crude SOD preparation (Figure 1). The reactivity
in resistant seedling was more compared to uninoculated control and uninoculated/inoculated
susceptible control seedlings (Table 1). Maximum absorbance
(0.51) was at 4 hrs post-inoculation. SOD reactivity analyzed in different
tissues of seedling revealed a maximum reactivity with absorbance of
In DIBA also PAb-MnSOD reacted strongly with the pure Mn-SOD compared to crude SOD preparation (Figure 3a). Mn-SOD reactivity was higher in resistant genotypes after inoculation compared to other samples (Figure 3b). DIBA was also used to detect the reactivity of PAb-MnSOD in different tissues of seedlings (Figure 3c). It was observed that the PAb-MnSOD reactivity was higher in root of inoculated resistant seedlings. Anti-Mn SOD antibody was used to detect antigens in western blot of pearl millet seedlings (Figure 4). A single immunoreactive band was present in resistant and susceptible pearl millet seedlings before and after inoculation. The molecular weight of this protein was 35 KDa, which is the molecular weight as detected previously for purified pearl millet Mn-SOD. The antibody did not recognize Cu/Zn-SOD from crude extract of resistant pearl millet seedlings. Using immunofluorescent technique, we also investigated the localization of Mn-SOD enzyme in different tissues of the resistant and susceptible pearl millet seedlings. Mn-SOD was most pronounced in vascular zone and hardly detected in the cortex (Figure 5). In the absence of secondary antibody bright yellowish auto fluorescence was observed in the root section. When the secondary antibody was added, green antibody-specific fluorescence was observed in the epidermal layer and the vascular bundle of root sections of the resistant seedlings. The green fluorescence was more pronounced in the root section after inoculation compared to the uninoculated root section. There was no remarkable difference in green antibody specific fluorescence in the shoot section of the resistant seedlings before and after inoculation (results not shown). On the contrary in susceptible seedlings also there not much difference in green fluorescence of both shoot and root sections before and after inoculation (data not shown). Our previous work demonstrated a clear differential induction of SOD activity in downy mildew resistant pearl millet seedlings after inoculation with S. graminicola (Babitha et al. 2002a). This is in agreement with the observations on several other pathosystems like potato tuber infected with Phytophthora infestans (Doke, 1985), Phaseolus vulgaris and Pseudomonas syringae (Croft et al. 1990) tobacco with blue mold pathogen Peronospora tabacina (Edreva et al. 1991) and coffee with Hemelia vastatrix (Daza et al. 1993). The protection of pearl millet against S. graminicola infection correlated with the level of increased SOD activity. The stepwise purification procedure for Mn-SOD consisted of ammonium sulphate precipitation, anion-exchange chromatography and gel-filtration on Sephadex G-100. Mn-SOD was purified 73-fold from pearl millet. Electrophoresis revealed a single band of SOD activity corresponding to the purified enzyme (Babitha et al. 2002b). This study determines the validity of utilizing the various immunological assays for determining Mn-SOD reactivity in pearl millet seedlings. To accomplish this Mn-SOD was purified from inoculated resistant pearl millet seedlings. The purified enzyme was utilized to develop sensitive and reproducible immunoassays, the specificity of which was tested by various serological tests. Polyclonal antibody against Mn-SOD has been developed from few sources like Paxillus involutus (Jacob et al. 2001), tobacco (Van Camp et al. 1996) and Arabidopsis (Kliebenstein et al. 1998). The present paper describes the attempts made and strategy employed to obtain specific polyclonal antibody against Mn-SOD and to develop a sensitive, rapid and specific ELISA to employ in routine downy mildew disease screening technique among the various pearl millet genotypes with varying degree of susceptibility/resistance. The antiserum was found to be extremely immunogenic as assessed by the titration of the immune sera. The ELISA developed was shown to be very sensitive with a titre of 1:20000. The antibody reacted more strongly with pure Mn-SOD than with crude SOD extract as shown by ELISA and DIBA. The SOD reactivity was significantly higher in inoculated pearl millet seedlings as compared to the uninoculated pearl millet resistant seedlings as detected by ELISA and DIBA. Tissue-specific expression as shown by ELISA and DIBA revealed a high SOD reactivity in inoculated root of resistant seedlings. The data from ELISA and DIBA suggest a correlation between SOD reactivity, SOD activity and degree of resistance of pearl millet genotype. To investigate the potential of immunological systems for identification of SOD, antiserum has been produced which shows specificity towards the antigen used but also confirms the existence of variation between genotypes of pearl millet. Additional serological tests like western blot and immunofluorescence confirmed the results. Western blots of pearl millet seedlings revealed a single band, corresponding to the 35 kDa purified protein. It showed an increased accumulation of SOD protein in the resistant seedlings after inoculation. Recombinantly expressed mitochondrial Mn-SOD was used to raise polyclonal antibodies, which cross-reacted with Mn-SOD in peroxisomes purified from pea leaves. Western blot assays of crude extracts with the antibodies to pea mitochondrial Mn-SOD showed that the levels of total Mn-SOD protein gradually increased with leaf senescence. Anti-SOD antiserum, when used in immunofluorescence experiments revealed two major locations for the antigen observable in fluorescence micrographs; the vascular bundle and the epidermis. The absence of specific fluorescence in the control tests indicated that the resultant staining depended only on antibody-antigen binding sites. EM immunochemistry was used to distinguish mitochondria and peroxisomal Mn-SOD in senescent leaves. Increased Mn-SOD labeling of perxisomes did not change with senescence (del Rio et al. 2003). The serological procedures used in these studies were sensitive enough to detect SOD in pearl millet genotypes. In summary, the data presented in this paper indicate that SOD can be detected immunologically in pearl millet and have sufficient specificity for simple identification purposes.
Note: Electronic Journal of Biotechnology is not responsible if on-line references cited on manuscripts are not available any more after the date of publication. © 2006 by Pontificia Universidad Católica de Valparaíso -- Chile The following images related to this document are available:Photo images[ej06016f4.jpg] [ej06016f3.jpg] [ej06016f1.jpg] [ej06016f2.jpg] [ej06016f5.jpg] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}