 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Electronic Journal of Biotechnology, Vol. 9, No. 2, April 15, 2006, pg. 144-151 RESEARCH ARTICLE Protective effect of an antimicrobial peptide from Mytilus edulis chilensis expressed in Nicotiana tabacum L. Gloria Arenas*1, Sergio H. Marshall2, Valeria Espinoza3, Ingrid Ramírez4, Hugo Peña-Cortés5 1Laboratorio
de Genética e Inmunología Molecular,
Instituto de Biología,
Pontificia Universidad
Católica de Valparaíso,
Av. Brasil 2950,
Valparaíso, Chile,
PO Box 4059,
Tel: 56 32 273205,
Fax: 56 32 596703,
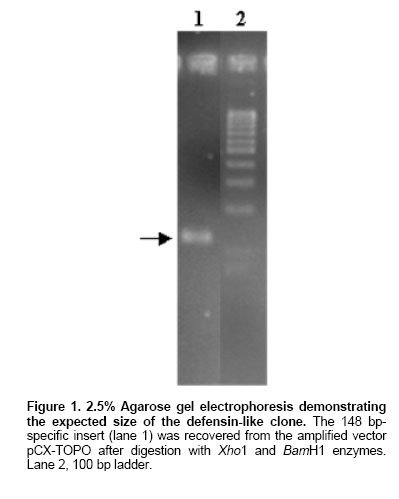
E-mail: garenas@ucv.cl Financial support: This work was supported by Grant PUCV-DI 122-769- Chile. Received August 2, 2005 / Accepted September 29, 2005 Code Number: ej06017 Abstract A “defensin-like” antibacterial peptide from Mytilus edulis chilensis, was sub-cloned into a binary vector for expression in plant tissues. The resulting new clone was electroporated into A. tumefaciens to transform tobacco plants. The presence of the construct in transgenic tobacco lines was demonstrated through RT-PCR, Northern and Western blots. Transformed positive plants were selected and grown for challenging. Tobacco leaves were infiltrated with Pseudomonassyringae pv. syringae and visual lesions determined at different times post-exposure. Of seven plants exposed, four gave variable protection up to seven days post-infection while one of them appears to be fully protected. These results suggest that defensin-like antimicrobial peptides from molluscs are a good source to provide resistance of tobacco plants to Pseudomonassyringae pv. syringae. Keywords: antibacterial peptide, disease resistance, in vivo expression, Nicotiana tabacum L., Pseudomonassyringae pv. syringae, transgenic tobacco plants. Antibacterial peptides and proteins are part of the innate response, and as such constitute the first line of defence against microbial infections in most living organisms (Zasloff, 2002; Yeaman, 2003; Tossi, 2005). They have been reported at all levels, from mammals (Ackermann et al. 2004; Bardan et al. 2004) to insects (Otvos Jr., 2000), marine invertebrates (Haug et al. 2004; Tincu and Taylor, 2004) and plants (Castro and Fontes, 2005). Most of these peptides have been successfully cloned, expressed and tested both in vitro and in vivo (Wu et al. 2000; Metlitskaia et al. 2004; Ingham et al. 2005) in their ability to neutralize bacterial infections. At present, more that 500 different molecules have been reported (http://aps.unmc.edu/AP/main.php). In mussels, the defensins constitute the most important peptide family characterized considering their primary structure and their consensus cysteine array (Charlet et al. 1996; Hubert et al. 1996; Mitta et al. 2000; Marshall and Arenas 2003). In plants, genes encoding defensins are developmentally regulated, with a predominant expression in outer cell layers (Moreno et al. 1994; Terras et al. 1995), and can be induced above basal levels in response to pathogen infection and other stresses (Chiang and Hadwiger, 1991; Moreno et al. 1994; Terras et al. 1995; Penninckx et al. 1996; Epple et al. 1997). Additionally, certain defensin genes are down-regulated by some pathogens (Moreno et al. 1994). Gene expression patterns of defensins (similar to plant thionines) are thus consistent with a hypothetical role in plant defence (Broekaert et al. 1995; Broekaert et al. 1997). There are a number of reports demonstrating the expression of heterologous antimicrobial peptides in plants with different degrees of success (Huang et al. 1997). The most abundant are those relating transferring these molecules from plant to plant (Sharma et al. 2000; Kanzaki et al. 2002). Notwithstanding, there are a few reporting transgenic expression of peptides derived from insect and amphibians (Sharma et al. 2000; Ponti et al. 2003). In this report, we demonstrate the protective expression of a defensin-like gene from Mytilus edulis chilensis as a stable transgene in Nicotiana tabacum plants when challenged with the phytopathogen Pseudomonassyringae pv. syringae. The 148
bp fragment was obtained from genomic DNA from Mytilus chilensis using
a set of degenerateprimers constructed upon the DNA sequence from Mytilus
galloprovincialis (Mitta et al. 1999). The pair of
primers which efficiently amplified the putative gene was: SEN-A, 5’-CCGCATCKATACCANGTRCANC- Construction of the expression vector The pCX-TOPO-containing clone was digested with Xho1 and BamH1 enzymes to release the defensin-like coding sequence. Prior to cloning the target sequence into the pBinAR plant tissue expression vector (Rocha-Sosa et al. 1989), the fragmentwas subcloned in the intermediate vector pBS in order to leave the target sequence in a “sense” orientation under the control of the 35 S CaMV promoter. pBS was linearized with Xho1 and BamH1, ligated with the defensin ORF, and positive clones detected through agarose gel electrophoresis. Restriction with Kpn1 -Xba1 enzymes released the coding sequence from the pBS vector which was exposed to the already Kpn1 -Xba1 linearized pBinAR vector. Ligation resulted in the recombinant expression vector. Positive clones (MCh-Def) were selected through standard bacterial transformation and restriction gel analysis as well as transformation and selection into Agrobacterium tumefaciens. Agrobacterium tumefaciens-mediated plant transformation The prepared construct MCh-Def was introduced in A. tumefaciens strain C58C1 containing plasmid pGV2260 (Deblaere et al. 1985) via electroporation. Transformation of tobacco (Nicotiana tabacum var. Xanhi NN) was performed using A. tumefaciens-directed gene transfer essentially as described (Fillatti et al. 1987). Presence of the defensin-like gene in transgenic plants Plants
in tissue culture were kept under a 16-h light/8-h dark period on Murashige
and Skoog medium (Murashige and Skoog, 1962) supplemented
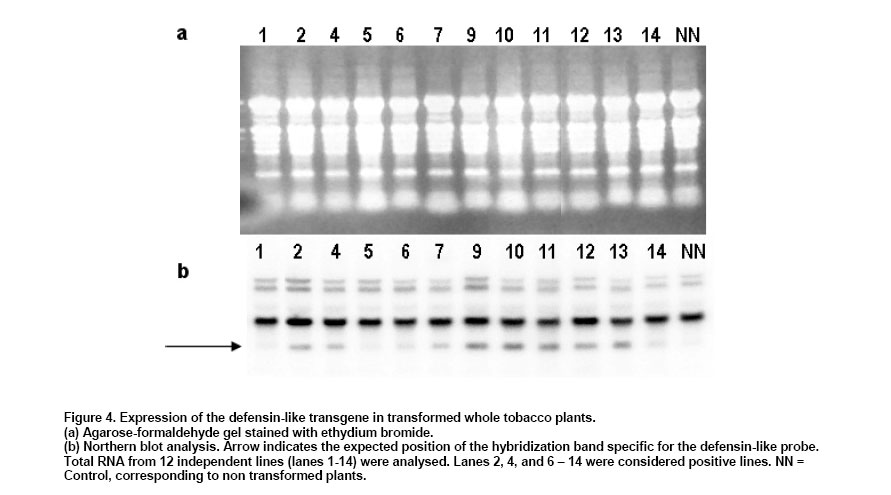
with 2% (w/v) sucrose at Expression of the defensin-like gene in transgenic plants Northern blots. Total RNA from control and transgenic tobacco plants were resolved in formaldehyde agarose gels and transferred to a nylon membrane (Harms et al. 1998). The membrane was hybridized to a 32P-labeled defensin-like probe. RT
PCR. The procedure described by Invitrogen SuperScript III First-Strand
Synthesis System for RT-PCR (Cat. No: 18080-051) was used. The usual precautions
for avoiding RNA degradation were taken until the reverse transcription
reaction was complete. A solution of 10 µg total RNA in 9 µl of 1X annealing
buffer was prepared and complemented with 1 µl of the oligodeoxynucleotide
degenerate primer mix (100 pmol/µl). The mix was heated for 3 min at Western blots. Total proteins from control and transgenic tobacco plants were determined by the bicinchoninic acid kit (Sigma Co. USA, B 9643), resolved in a 10% Laemmli PAGE-SDS gel, and transferred to a nitrocellulose membrane. The membrane was then exposed to a 1/5000 dilution of the polyclonal antibody MGD1, elicited in rabbits against a Mytilus defensin (Mitta et al. 1999) as the first antibody followed by an incubation with an HRP-labelled anti rabbit IgG fraction and developed with DAB (Towbin et al. 1979). Challenge with Pseudomonas syringae pv. syringae Leaves
from transgenic and control tobacco were infiltrated by mechanical injury
with the phytopathogen Pseudomonas syringaepv. syringaeat a
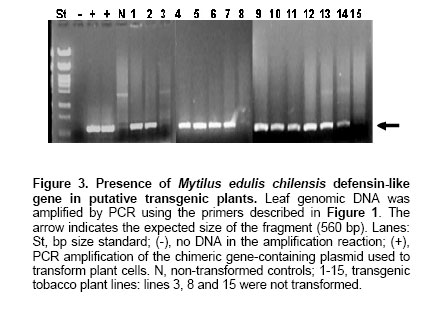
concentration of approximately 105-106 CFU/ml in Generation of the MCh-Def clone Figure 1 shows the DNA insert corresponding to the defensin-like ORF from Mytilus edulis chilensis. This was subcloned into the pBinAR plant tissue expression vector as shown in Figure 2. Detection of the defensin-like transgene in transformation-positive tobacco plants PCR. As
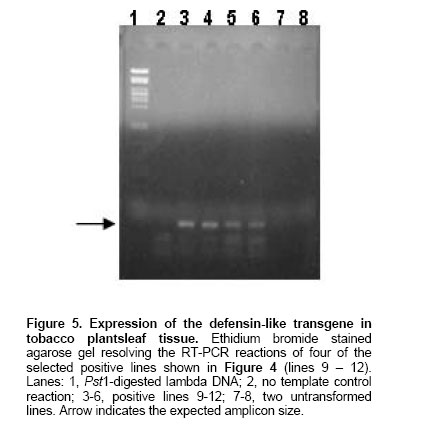
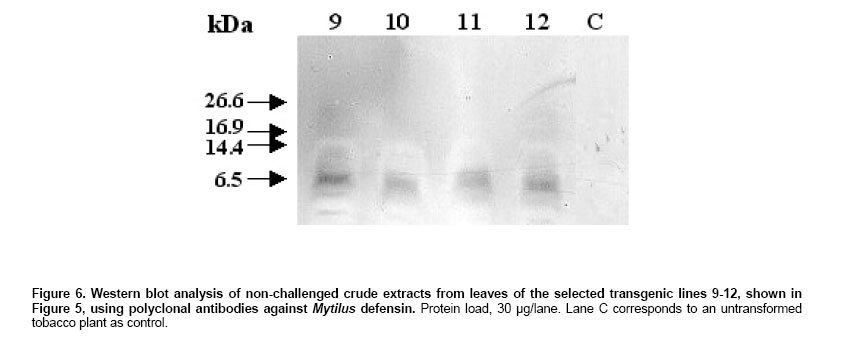
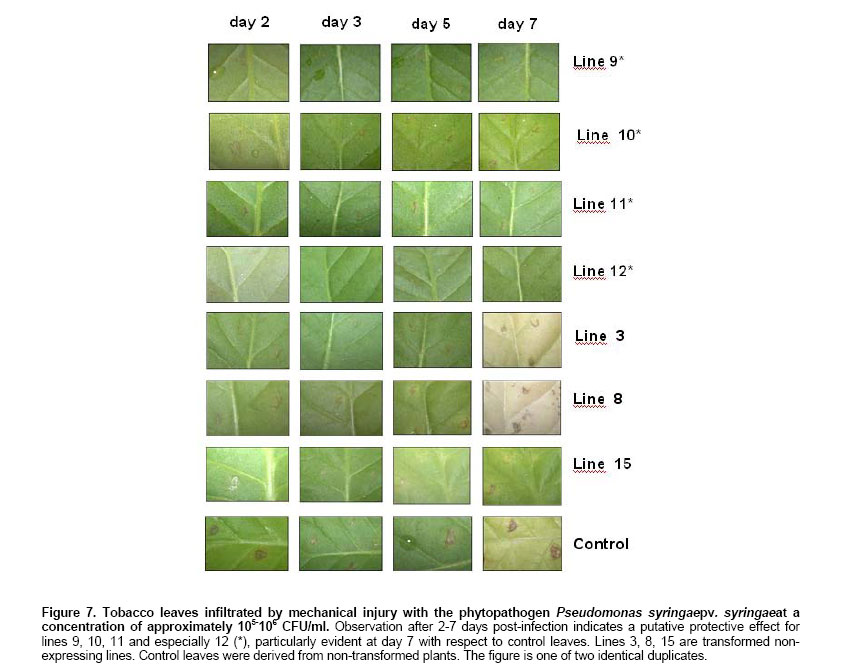
shown in Figure 2 Northern Blot. Selected lines, grown under axenic conditions, were the source for total RNA extraction in order to detect the putative transcript from the defensin-like transgene. Figure 4a shows the stained profile of the material resolved on the agarose gel, while Figure 3 shows the result of the transferred material hybridized to the defensin-like specific probe. Ten out of the 12 selected lines gave different degree of hybridization to the probe indicating differential transcription in the lines which correspond to the expression of MCh-Def gene (lanes 2, 4 and 6-14, respectively). RT PCR. In order to reconfirm the results from Northern blot analysis, four out of the seven transcription positive lines were selected to evaluate specific transcription on leaf tissue which will be the target of challenging with pathogen bacteria. Figure 5 shows the RT-PCR results, confirming that lines 9-12 do express the defensin-like transgene. Western Blot. Further, in order to detect recombinant peptide in the selected lines, crude protein extracts of leaves were resolved on denaturant gels, transferred to a nitrocellulose membrane and exposed to a defensin-like specific antibody (Figure 6). Challenge with Pseudomonas pv. syringae and evaluation of resistance Transgenic as well as control tobacco leaves were infected with P. syringae to evaluate the degree of resistance provided by the expressed defensin-like transgene. Figure 7 shows the degree of damage observed. There have been a number of attempts to use both heterologous and even homologous peptides as tools to increase disease-resistance in transgenic plants (for a review, see Castro and Fontes, 2005). Although most of them have been unsuccessful,some of them have been inspiring in the search for new alternatives (Sharma et al. 2000; Kanzaki et al. 2002; Ponti et al. 2003). Some of the reasons for the lack of success in the past have been estimated to be due to either a low level of expression of the transgene (Halpin, 2005), a putative inactivation of the gene product by constitutive proteases in the new host environment, or a low half-life of the expressed peptide in the host plant (Owens and Heutte, 1997). In this report, the use of a vector containing the defensin-like ORF from Mytilus edulis chilensis in frame with the 35S promoter from the cauliflower mosaic virus (Figure 2) allowed us to get an 80% efficiency of transformation and a reasonable good expression which offers a detectable, protection to tobacco leaves when challenged with Pseudomonas syringae pv. syringae. We selected 12 positive transgene lines containing the M. edulis chilensis defensin-like cDNA to demonstrate disease-resistance (Figure 3). Notwithstanding not all tissues of the positive lines either had the transgene and/or express it. Northern blot analysis indicated that only 7 out of 12 independent transgenic lines gave a significant expression and one of them, a faint one (Figure 4, lanes 2, 4, and 9-13). We selected lines 9-12 to further verify RNA expression in leaves of the transgenic plants, the target of our challenging test. Indeed all four positive lines gave significant transcription of the transgene (Figure 5). Additionally, the protein was expressed in all four lines (Figure 6), providing a strong argument to support the resistance to infection shown by challenged leaves measured by two unequivocal parameters: number and/or size of the lesions observed (Figure 7). In this frame, protective responses were detected in the positive lines up to 7 days post-challenge, well above to those described for other peptides in previous works on tobacco plants (Okamoto et al. 1998; Koo et al. 2002; Li et al. 2001). These positive preliminary results are encouraging. Considering that the transgene product is a defensin-like molecule expressed in a system which has natural counterparts (Castro and Fontes, 2005), endogenous proteases do not consider it a target. In addition, the transgene product could act synergistically with endogenous AMPs, natural defensins and/or alike, which might explain the observed protective effect over challenging in selected transgenic lines. Because endogenous defensins are expressed in minute amounts in tobacco leaves, over-expression of the transgene, even under non-challenged conditions, transforms it in a helper-like tool to overcome putative encounters with aggressive pathogens. In conclusion, this report is a contribution to further design strategies to evaluate the impact of family-related peptides in commercially relevant plant species which might be optimized through their enhanced disease-resistance potential.
Note: Electronic Journal of Biotechnology is not responsible if on-line references cited on manuscripts are not available any more after the date of publication. © 2006 by Pontificia Universidad Católica de Valparaíso -- Chile The following images related to this document are available:Photo images[ej06017f3.jpg] [ej06017f7.jpg] [ej06017f4.jpg] [ej06017f6.jpg] [ej06017f2.jpg] [ej06017f1.jpg] [ej06017f5.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}