 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Electronic Journal of Biotechnology, Vol. 9, No. 5, October, 2006, pg. 559-565 RESEARCH ARTICLE Immobilization and stability studies of a lipase from thermophilic Bacillus sp: The effect of process parameters on immobilization of enzyme Neerupma Nawani1, Rajvinder Singh2, Jagdeep Kaur*3 1Department of Biotechnology,
*Corresponding author Financial
support: Junior Research Fellowship to N.N. from Council of Scientific and Industrial Research,
Received December 2, 2005 / Accepted May 3, 2006 Code Number: ej06075 Abstract A thermostable lipase was partially purified from the culture supernatant of a thermophilic Bacillus sp. The enzyme is optimally active at
Keywords: esterification, immobilization, lipase, thermostability. Lipases, as a class of enzymes, are stable and extremely valuable catalysts for many practical/industrial applications (Bjorkling et al. 1991). They have been used to generate chiral entities from alcohols, carboxylic acid esters, cyanohydrins, chlorohydrins, diols, amines, diamines and amino alcohols (Jaeger et al. 1999), which are used as building blocks for a variety of pharmaceuticals and other fine chemicals. Many lipases are only moderately stable at high temperature and pHs, that can influence their usefulness in some interesting reactions. Using lipases from thermophilic microorganisms, whose resistance to drastic conditions has been developed by nature, can solve this problem. At present, the majority of the thermophilic lipases that have been purified and characterized are obtained from Bacillus sp (Kim et al. 1994). To use these enzymes for industrial purposes, the most desirable step is to improve the features of biological catalyst to suit the industrial demand. The majority of current biocatalytic approaches rely on either using free enzyme in solution or living cells, which complicates product separation from the catalyst. The lipase immobilization may improve the stability and ease of product separation/reuse of enzyme. Although a variety of approaches exist for the immobilization of enzymes, the "science" of enzyme immobilization is still in its infancy. Several solid supports have been used for the purpose till now (Aucoin et al. 2004; Hwang et al. 2004; Palomo et al. 2004). A novel thermophilic Bacillus sp has been isolated that produced a thermostable lipase from
The culture (isolated Bacillus sp) was grown under optimal conditions for lipase production. A series of 500 ml, Erlenmeyer flasks containing 100 ml wheat bran and nutrient broth medium, (1% w/v each) pH 8.0 were seeded with 2% inoculum and incubated at
The activity was determined according to the method of Sigurgisladottir et al. (1993) with slight modification. To 0.8 ml of
Partial purification of enzyme Ammonium sulphate precipitation. To 900 ml of the culture supernatant, ammonium sulphate was added (70% saturation) at
Hydrophobic interaction chromatography. The precipitated enzyme (276 ml) was loaded on Phenyl Sepharose (Sigma) column (4.0 x
Effect of organic solvents on enzyme activity To study the effect of organic solvents on aqueous enzyme activity, organic solvents such as methanol, ethanol, acetone, Dimethylsulphoxide (DMSO), glycerol, polyethyleneglycol, hexane, benzene (10%, 30% v/v) were added to the reaction mixture and the lipase activity was determined under standard assay conditions. The sample without any additive was taken as control (100%). The reaction mix with respective additives but without enzyme served as control. In addition enzyme was assayed in the presence of 60% of hexane or benzene in reaction mixture. Adsorption kinetics of protein and lipase activity on solid support. Silica (Merck) and HP-20 beads (Diaion) were used for immobilization. HP-20 is aromatic in chemical structure (surface area ca. 600 m2/g, pore radius 200-
Silica. 5 ml of partially purified enzyme was mixed with
HP-20 beads. 1g of HP-20 was first washed with distilled water, then washed for 30 min with isopropyl alcohol and further washed three times with distilled water to remove isopropyl alcohol traces. The half of the washed beads were used as such and rest half was treated with 2.5% gluteraldehyde for 1 hr. The resin was washed 3 times with distilled water to remove residual gluteraldehyde. 5 ml of enzyme was added to the resin and left on shaker for 90 min. After different time points (0-90 min) fractions were taken out and centrifuged at
 Adsorbed lipases on different solid supports were assayed to define the adsorption capacity. Hundred mg of silica and HP-20 support (with and without cross-linking) with immobilized lipase was added into assay mixture and assayed with continuous shaking for 30 min as per standard method. The enzyme activity bound on each support was determined. Retention of activity by immobilized lipase in continuous cycles. The immobilized enzymes were assayed by the method of Sigurgisladottir et al. (1993) for 25 cycles of 30 min each. For each cycle two ml of reaction mixture containing substrate was added to the immobilized support and incubated for 30 min with continuous shaking at
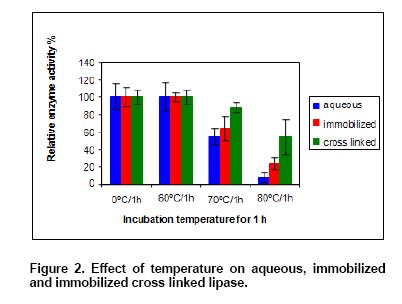
Thermo-inactivation of free and immobilized lipase. To study the effect of temperature on free, immobilized and immobilized cross-linked lipase, different forms of enzymes were incubated at different temperatures (60, 70 and
To determine the half life of lipase, free (80 U/ml) and immobilized (180 U/g) lipase was incubated at
Effects of pH and temperature on enzyme activity. Lipase activity was assayed in the buffers of different pH (4.5-11.0) at
The optimum temperature of lipase activity was determined by carrying out the enzyme reactions at different temperatures (30-
Esterification studies by aqueous and immobilized lipase For comparative study, aqueous and immobilized enzymes were used as biocatalysts for the esterification of oleic acid (
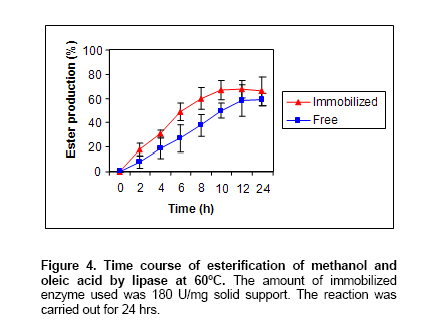
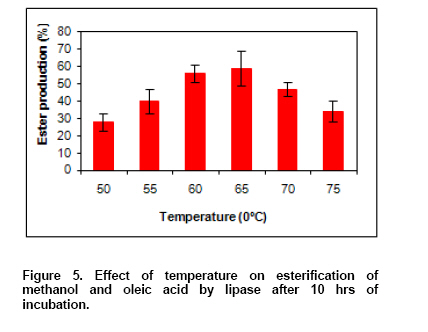
Time course of esterification. To determine the optimum time for the formation of methyl oleate from methanol and oleic acid by immobilized enzyme, the esterification reaction was carried out for different time points. The ester synthesis was analyzed as above. Effect of temperature on esterification reaction. The optimum temperature for the formation of methyl oleate from methanol and oleic acid by immobilized enzyme, the esterification reaction was carried out at different temperatures (50-
Basic characteristics of free lipase The strain was identified as Bacillus sp. based on characteristics such as aerobic growth, gram positive, rod shaped, motile, spore forming and catalase positive (Clans and Berkley, 1986). The organism could hydrolyze starch, casein and gelatin. It did not require sodium chloride and potassium chloride for growth. It was oxidase negative in nature and it did not produce any gas with glucose. On comparison of this Bacillus sp isolate with Bacillus stearothermophilus and other thermophilic Bacillus sp. (according to Bergey's manual), it showed the following different characteristics i.e. nitrate reduction, acid formation with arabinose, xylose and mannitol, casein hydrolysis, and no growth below
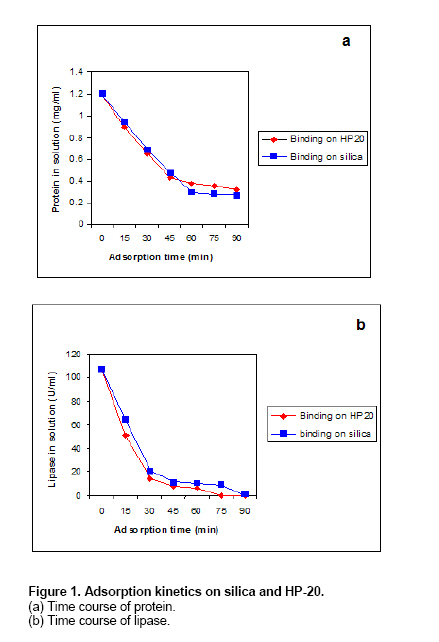
The lipase was partially purified by ammonium sulphate precipitation followed by Phenyl-Sepharose column chromatography (specific activity 160 U/mg protein). This partially purified enzyme was used for immobilization. As to the effect of organic solvents on enzyme activity, benzene and hexane had a highly stimulatory effect on the lipase activity (195 and 180% respectively). The activity of enzyme was stimulated up to 60% of hexane/benzene in reaction mixture. The enzyme activity was slightly stimulated up to 30% in presence of acetone. DMSO (Dimethylsulphoxide) and ethylene glycol had almost no effect, while ethanol, methanol and propanol inhibited the enzyme activity. The effect of organic solvents might be attributed to the water content in reaction mix. Water plays a significant role in the activity of the enzymes (Klibanov, 1989). The inhibition of lipase in presence of ethanol, methanol while stimulation in water immiscible solvents confirms that polar water miscible solvents are more destabilizing than the water immiscible solvents (Fernández-Lorente et al. 2001). On the other hand effect of acetone on enzyme activity could not be explained by this theory. The immobilization by ionic or physical adsorption was one of the simple and cheap methods to prepare an immobilized enzyme, although adsorbed enzymes were rather susceptible to desorption from the carrier (Rua and Ballesteros, 1994). Immobilization of isolated Bacillus sp lipase was carried out on HP-20 and Silica. The adsorption of lipase on various supports was determined by estimating the amount of lipase bound to the solid supports (Table 1). It was found that good adsorption was seen in HP 20 (with and without cross linking) and silica, where attachment to lipase was up to the extent of 78, 79 and 86% respectively. The kinetics of protein and lipase adsorption to silica and HP-20 indicate that the amount of protein and lipase was well below saturation (Figure 1a) while no unbound lipase could be traced (Figure 1b). The protein and lipase adsorption was fast and maximum binding was observed after 45 min for protein and 30 min for lipase. This suggests that lipase is fast adsorbing protein amongst other proteins. The hydrophobic nature of solid substrate suggests that adsorption of proteins are governed by hydrophobic interactions. Rua and Ballesteros (1994) used moderately hydrophobic support (Phenyl Agarose) in the purification of lipase from C. rugosa. Similar hydrophobic adsorption supports were used by other workers for immobilization (Aucoin et al. 2004; Palomo et al. 2004). Continuous assay of residual enzyme activity in the lipase immobilized to HP-20 and silica was performed to find out the retention of lipase activity by each support over 25 cycles of enzyme reaction. Less desorption or leaching of the bound enzyme in case of HP-20 cross-linked might be due to very tight binding of the lipase on the hydrophobic supports, (Table 2). The lipase immobilized on HP-20 without cross-linking behaved like silica (data not shown) and in both the cases the leaching of enzyme activity was observed. Lipase immobilized on silica might be lost from the system because of de-sorption, severing of chemical bonds or erosion of the support material. For clear presentation of results only enzyme immobilized on silica is compared with HP-20 cross-linked. Similar to our observations, Fernández-Lorente et al. (2001) could use the immobilized lipase using hydrophobic moieties for 10 esterification cycles without any significant decrease in enzyme biocatalyst.
Physico-chemical characterization and thermal stability of immobilized lipase No change in optimum pH for lipase activity was observed in the case of aqueous and immobilized form. Optimum temperature of immobilized (CL) enzyme shifted from
Half-life of aqueous and immobilized lipase (HP-20) was calculated to be 85-90 min and 4 hrs respectively at
The biosynthesis of esters is currently of much commercial interest because of the increasing popularity and demand for natural products amongst consumer. Bio-transformations and enzymatic methods of ester synthesis are more effective when performed in non-aqueous media (Chand et al. 1997). Because of the fact that the present lipases showed much activity and stability in hexane,the esterification of oleic acids and methanol in hexane was carried out. The effect of incubation time on esterification reaction indicates that ester synthesis was essentially complete by 9-10 hrs with immobilized enzyme while it took more than 12 hrs with aqueous enzyme (Figure 4). No enhancement in ester production was observed after 10 hrs. Fig 5 illustrates the effect of temperature on esterification reaction. During the esterification reaction, water is released as a byproduct and was to be inhibitory to the esterification reaction at a particular concentration (Chand et al. 1997). The higher conversion rate observed in the case isolated lipases here may be explained by the reduction of the water content due to the higher incubation temperature thus decreasing the inhibitory effect of water on esterification reaction. In esterification conditions, one molecule of water is produced when one molecule of acid is esterified. In order to favor this reaction, the produced water must be removed from the medium. Selmi et al. (1997) described temperature increase with air ventilation as one of the possibility. The degree of esterification increased with temperature in the range of (50-
For any application based on immobilized lipases, the feasibility of regeneration of the lipase activity (and consequent reuse of the support) provided clear economic benefits for its industrial use. HP-20 beads were comparatively cheap and quite rigid in nature. It had been demonstrated that the immobilization and cross linking of the lipase on HP-20 was advantageous for catalytic property and stability
Note: Electronic Journal of Biotechnology is not responsible if on-line references cited on manuscripts are not available any more after the date of publication. © 2006 by Pontificia Universidad Católica de Valparaíso -- Chile The following images related to this document are available:Photo images[ej06075f2.jpg] [ej06075f3.jpg] [ej06075f1.jpg] [ej06075f5.jpg] [ej06075f4.jpg] | ||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}