 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Electronic Journal of Biotechnology, Vol. 9, No. 5, October, 2006, pg. 572-579 RESEARCH ARTICLE Multiple gene knock-down by a single lentiviral vector expressing an array of short hairpin RNAs Veronique Stove1 , Kaatje Smits2 , Evelien Naessens3 , Jean Plum4 , Bruno Verhasselt*5 1Department of Clinical Chemistry,
Microbiology and Immunology,
Financial support: This work was supported by grants from the Research Foundation – Flanders (FWO). V.S. and K.S. equally contributed and are Ph.D. fellows of the FWO, and B.V. is a Senior Clinical Investigator of the FWO. Received December 28, 2005 / Accepted May 29, 2006 Code Number: ej06077 Abstract RNA interference (RNAi), mediated by short double-stranded RNAs, is a powerful mechanism for posttranscriptional gene silencing. Sustained expression of short hairpin RNA (shRNA) can be accomplished in mammalian cells by viral delivery systems. Using lentiviral constructs, stable gene silencing is established both in dividing and non-dividing cells. Targeting one single gene can lead to the development of escape mutants or may be insufficient to silence redundant pathways. Therefore, simultaneous targeting of multiple genes may be necessary. We have generated a lentiviral vector-based system for expression of multiple shRNAs from a single viral vector, which also encodes an EGFP reporter protein. We show that knock-down of each single gene from multiple target vectors is achieved at an efficiency comparable to that obtained after transduction using single target viral vectors. In this way, we were able to knock-down several members of the human Rho-family GTPases in T cells. Double and triple knock-down persisted after multiple passages of the cells. The ability to inhibit two or more genes simultaneously from one single expression vector further widens the application spectrum of RNAi, both in functional studies and therapeutic strategies. Keywords: EGFP, lentiviral gene transfer, multiple knock-down, RNAi, Rho GTPases, shRNA.
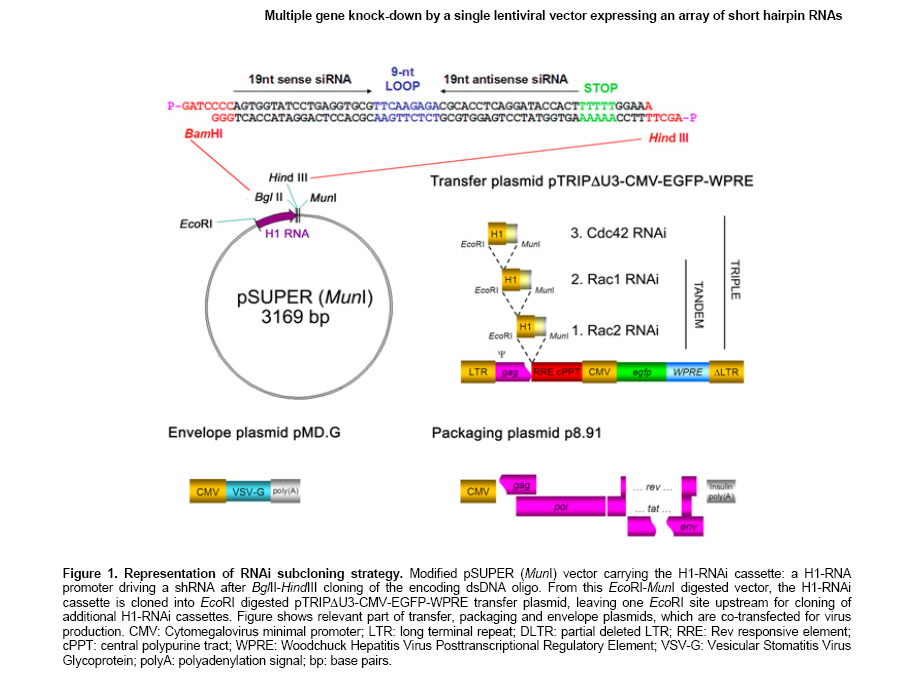
Knock-down by small interfering RNAs (siRNAs) has been shown to be a powerful technique. Transfer of siRNA to cells can be achieved in several ways, e.g. delivered exogenously or expressed endogenously from RNA polymerase III (pol III) promoters (reviewed by Dykxhoorn et al. 2003). Initial reports using siRNA in mammalian cells focused on the transient knock-down of target gene expression, using direct transfection of synthetic siRNA, or transient expression of double stranded RNA via transfection of plasmid DNA, bearing a RNA pol III promoter-driven expression cassette. Several reports have also shown the feasibility to knock down several proteins simultaneously (Anderson et al. 2003; Yu et al. 2003; Fitzgerald et al. 2004; Lakka et al. 2004). Moreover, the combination of siRNAs directed against different regions of a single gene can be used to improve the gene silencing effect or to avoid viral escape mutants (Ji et al. 2003; Gonzalez et al. 2005; Schubert et al. 2005). Multiple RNAi can be accomplished by transfection of multiple siRNA species (Ji et al. 2003; Yu et al. 2003). Alternatively, a construct with several consecutive short hairpin RNAs (shRNAs) expression cassettes (Gonzalez et al. 2005; Jazag et al. 2005; Schubert et al. 2005), or expression of a RNA that is cleaved into several shRNAs, are an option (Anderson et al. 2003; Lakka et al. 2004). For expression in primary cells or cell lines difficult to transfect, only viral transfer reaches a significant efficiency (Verma and Somia, 1997). More specifically, lentiviral vectors have been shown to be a valuable approach for stable and effective transfer of functional shRNAs in these cells (Qin et al. 2003; Rubinson et al. 2003). To extend the application possibilities of lentiviral vector mediated knock-down, we developed lentiviral vectors encoding within one construct distinct shRNAs, each driven by an individual pol III promoter. This ensures simultaneous expression of all distinct shRNAs. We show that these multiple shRNAs vectors down-modulate the respective targets at a comparable level as the single target vectors. As proof of principle we used these vectors to silence the expression of up to three In this study, we have examined the functionality of different shRNA cassettes, inserted one after the other in a single lentiviral construct. We found that these recombinant lentiviruses were able to stably knock down the expression of the targeted genes on mRNA level as well as on protein level. The pSUPER plasmid containing H1 RNA pol III promoter, described previously (Brummelkamp et al. 2002), was obtained from Oligoengine (Seattle, WA). A unique MunI site was inserted downstream the HindIII restriction site, creating the pSUPER(MunI) vector. The RNAi probes were BglII-HindIII cloned into this vector. To construct the 19 nucleotide hairpin siRNA cassettes, two complementary DNA oligonucleotides were chemically synthesized (Invitrogen, Merelbeke, Belgium), annealed, and inserted immediately downstream of the H1 promoter: 5'-GATCCCC-19-TTCAAGAGA-19-TTTTTGGAAA-
Lentiviral production was done as described before (Stove et al. 2005) using packaging plasmid p8.91 (Zufferey et al. 1997), vesicular stomatitis virus (VSV) envelope-expression plasmid (pMD.G) (Naldini et al. 1996) and TRIPΔU3-CMV-EGFP-WPRE transfer vector. As MunI and EcoRI generate compatible sticky ends, the H1-RNAi cassettes could be EcoRI-MunI transferred from the pSUPER(MunI) vector into the EcoRI digested TRIPΔU3-CMV-EGFP-WPRE vector. The EcoRI remains a unique restriction site after ligation and can still be used to subclone additional H1-RNAi cassettes into the lentiviral vector (Figure 1). Cell culture and viral production All the cells (SupT1 (AIDS Research and Reference Reagent Program, NIH, Bethesda, MD) and 293T (American Type Culture Collection, Rockville, MD)) were grown in Iscove's modified Dulbecco's medium (IMDM, Invitrogen) supplemented with penicillin (100 IU/ml, Invitrogen), streptomycin (100 µg/ml, Invitrogen) and 10% heat-inactivated fetal calf serum (PerBio,Erembodegem, Belgium). VSV-glycoprotein pseudotyped lentiviral vector particles were produced by co-transfection of 293T cells with transfer, packaging and envelope plasmids (respectively 20 µg, 20 µg and 10 µg / 175 cm² culture flask) using a Calcium Phosphate Transfection Kit (Invitrogen) and viral sup was harvested 40 hrs later. Viral supernatants contained at least 108 transducing units per ml titrated on SupT1 cells. Transductions were carried out in the presence of polybrene (4 μg/ml; Sigma-Aldrich, Bornem, Belgium) with multiplicity of infection (MOI) well above 1. Chemotaxis assays were performed in duplicate using 5 µm pore filters (Transwell, 24 well cell cluster, Corning Costar,
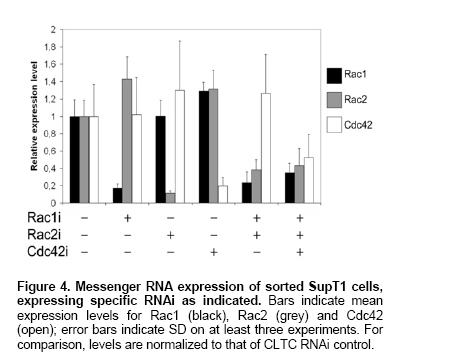
Upper wells were removed and cells migrated into the lower compartment were harvested after addition of a fixed amount of Flow-Count Fluorospheres (Beckman Coulter, Fullerton, CA). Flow cytometry was done on a FACS®Calibur (Becton Dickinson Immunocytometry Systems, Mountain View, CA) to determine the absolute number of input and migrated cells as well as the relative frequency of EGFP+ cells in initial and migrated population. Percentage of migrating cells was calculated as follows: (%EGFP+ cells in migrated population x total amount of migrated cells) / (%EGFP+ cells in initial population x total amount of input cells). Total RNA was extracted from sorted shRNA transduced cells as instructed by supplier (TRIzol, Invitrogen), DNase-treated (DNase I, Invitrogen) and reverse transcribed (Reverse Transcription Core Kit, Eurogentec, Seraing, Belgium). Genes assayed included Rac1, Rac2 and Cdc42 (TaqMan detection chemistry, Eurogentec) and β2-microglobulin housekeeping gene (B2M) (Sybr Green I detection, Eurogentec). Primers (Invitrogen) and Taqman probes (Eurogentec) were described previously (Kovacic et al. 2001; Vandesompele et al. 2002) or were designed with Primer Express 2.0 software (Applied Biosystems, Foster City, CA). Sequences were as follows (
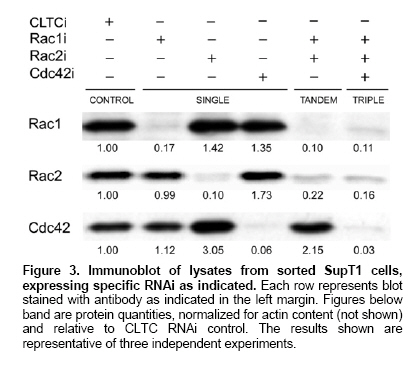
Sorted transduced cells were lysed in Laemmli sample buffer (
Lentiviral delivery of shRNAs targeted to a single
Several shRNAs - 2 shRNAs per targeted gene - were synthesized corresponding to the genes Rac1, Rac2 and Cdc42. Each shRNA also served as negative control for the two other, non-targeted genes. Furthermore, a shRNA targeting clathrin heavy chain (CLTC) was chosen as an additional control. To investigate the potency of the selected short hairpins to stably knock down their respective target, we designed lentiviral constructs expressing a shRNA from the pol III promoter H1 and an enhanced green fluorescent protein (EGFP) reporter from a CMV internal promoter. Both directions were possible for insertion of the siRNA cassette with equal capacity to knock down the target gene (data not shown). For simplicity, all the constructs were built with the internal promoters 5'-
Simultaneous knock-down of 2 or 3
To test the effectiveness of simultaneous knock-down of multiple genes, we inserted 2, and in a next step 3 RNAi cassettes 5'-
Functional implications of knock-down of
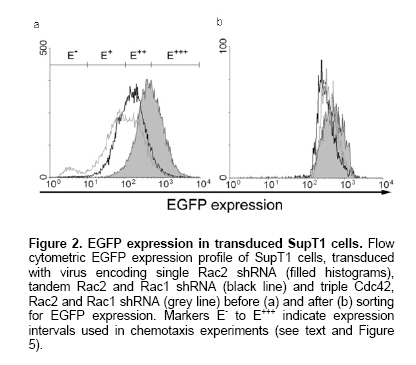
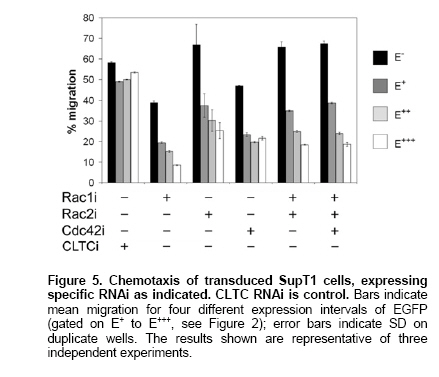
Rho GTPases are crucial proteins for directed cell migration (Raftopoulou and Hall, 2004). To investigate the functional correlate of the observed knock-down, we performed migration assays with SupT1 cells, known to respond chemotactically to SDF-1α (Secchiero et al. 1998). In these experiments, RNAi transduced cells were not sorted, allowing us to score migration in function of EGFP expression (gating on intensity regions as shown in Figure 2a, E- to E+++). As shown in Figure 5, knock-down of Rac1, Rac2 and Cdc42 all resulted in impaired chemotaxis towards SDF-1α. Inhibition of migration correlated with EGFP levels, indicating a biological read-out of the shRNA levels. Cells expressing shRNA migrated to a lower extent compared to cells not expressing shRNA when either one of the three Rho GTPases was knocked down, suggesting that targeted Rho GTPase proteins are all necessary for directional migration. Consequently, as expected from knock-down observed by immunoblot, this biological effect was retained in cells transduced with virus encoding multiple shRNAs (Figure 5). In this report, we generated lentiviral constructs to study knock-down of Rho GTPases, important in cell motility. Simultaneous knock-down of several individual Rho GTPases was achieved with multiple siRNAs, each driven by individual H1 pol III promoters, expressed from a single lentiviral vector. Thanks to a separate marker gene insert, the dose-response on knock-down can be monitored. In order to establish stable silencing of Rho GTPases, lentiviral constructs expressing shRNA were designed. Lentiviral gene transfer has been shown to be an efficient and gentle method to transfer siRNA into primary cells and by virtue of a marker gene, transduced cells effectively expressing shRNA can be identified. For effective knock-down of signaling cascades or redundant protein pathways, several targets have to be silenced in the same cell. In this study, we further extended the use of lentiviral siRNA transgenesis by generating tandem and triple constructs to achieve multiple protein knock-down. These constructs have an advantage over simultaneous transfection of several siRNAs, as the latter method does not ensure that all siRNAs are expressed simultaneously. Alternatively, multiple transductions with separate viruses, each encoding one shRNA, are feasible if high transduction efficiencies are attained. However, unless each vector encodes a different marker gene, it remains unclear if a single cell expresses all shRNAs intended to be transferred. Moreover, expression from a single vector has the advantage that the relative level of expression of each shRNA is constant. A method using plasmids expressing multiple siRNAs driven by U6 promoters to achieve knock down of escape mutants of a single target (Schubert et al. 2005) or to target different mRNAs (Jazag et al. 2005) were described recently. These and our results indicate that no apparent competition exists between multiple pol III promoters in close proximity and that no saturation of the RNA-induced silencing complex RISC was reached, as suggested previously (McManus et al. 2002). Moreover, Gonzalez et al. 2005 have recently reported a proportional improvement in HLA gene silencing by increasing the copy number of a selected U6-shRNA cassette in the plasmid vector from 1 to 6. Further increasing the number of RNAi cassettes showed a decrease in HLA down-regulation, evidencing a certain limit (competition or saturation) in shRNA expression capacity. However, this limit might be different depending on the targeted gene and the cell line or vector system that is used. Recently, reports have shown that simultaneous silencing of two different cellular targets is achievable using a single lentiviral vector containing two pol III promoters (Anderson and Akkina, 2005; Sumimoto et al. 2006). Our data corroborate these results and provide evidence that knock-down of three different targets from one type of pol III promoter is still feasible. Moreover, our design allows an easy gradual extension of the number of RNAi cassettes in the widely used lentiviral vector pTRIP, which can, based on its theoretical incorporation capacity (Verma and Somia, 1997), accommodate more than 10 shRNA cassettes. In this context of multiple identical promoter sequences in a row, the possibility of repeat sequence-mediated deletion during reverse transcription should be considered. However, we hypothesize that the net effect of this phenomenon might be minimal when using high MOI. The evaluation of other and more sets of shRNAs is warranted and currently in progress. Recently, nonspecific effects of siRNAs on gene expression have been reported (reviewed by Dykxhoorn and Lieberman, 2005). In our experiments, we did not observe ‘off-target' stimulation or repression for the investigated genes. Some protein quantification results showed higher protein levels of non-targeted Rho GTPases. However, this effect was not observed on mRNA level. Moreover, Rac1 could not compensate for loss of function of Rac2 and vice versa in the chemotaxis assay. Interestingly, inhibition of migration correlated with EGFP levels, demonstrating that the marker gene expression, although driven by a separate promoter, is a reliable measure for shRNA levels. This validates our approach of sorting similar EGFP intensities in transduced cells for direct comparison of the effect of shRNA expression. Moreover, the potential caveat of comparing different vector stocks with different MOI is avoided by this approach. Further functional experiments are needed to delineate the Rho GTPase network response to knock-down of individual members using our vectors. In conclusion, we present here a lentiviral shRNA delivery system that allows multiple shRNA species and a marker gene to be expressed from the same vector. The simultaneous knock-down of several targets within the same cell may be necessary, e.g. in compensatory or redundant pathways that have to be blocked at several levels, or to target hyper-variable viral sequences. Therefore, this strategy will likely prove to be of value in experimental and therapeutic applications. The authors thank Nancy De Cabooter and Els Demecheleer for technical assistance, Didier Trono (Université de Genève,
Note: Electronic Journal of Biotechnology is not responsible if on-line references cited on manuscripts are not available any more after the date of publication. Supported by UNESCO / MIRCEN network © 2006 by Pontificia Universidad Católica de Valparaíso -- Chile The following images related to this document are available:Photo images[ej06077f5.jpg] [ej06077f1.jpg] [ej06077f3.jpg] [ej06077f2.jpg] [ej06077f4.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}