 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Electronic Journal of Biotechnology, Vol. 9, No. 3, June, 2006, Supp. 1, 2006, pg. 199-204 RESEARCH ARTICLE Analysis of the molecular basis of Xanthomonas axonopodis pv. citri pathogenesis in Citrus limon Florencia Siciliano1 , Pablo Torres2 , Lorena Sendín3 , Carolina Bermejo4 , Paula Filippone5 , Gabriel Vellice6 , Jackie Ramallo7 , Atilio Castagnaro8 , Adrian Vojnov9 , María Rosa Marano*10 1Instituto de Biología Molecular y Celular de Rosario,
FByF, UNR, Suipacha 531, 2000,
Rosario, Argentina
Tel: 0341 435 6369
Fax: 0341 439-0465
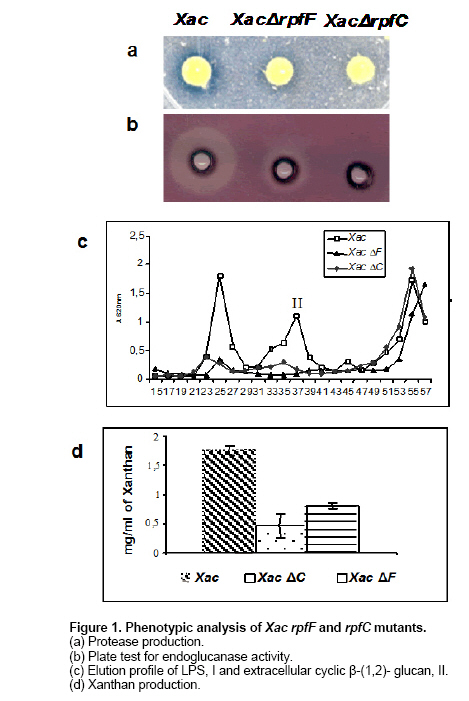
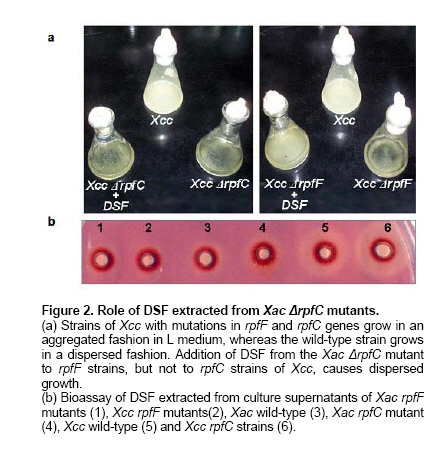
E-mail: florisiciliano@yahoo.com.ar Financial support: Agencia de Promoción Científica y Tecnológica (PICT-02 No: 08-10740). Code Number: ej06086 Abstract Xanthomonas axonopodis pathovar citri (Xac) causes bacterial citrus canker, a serious disease of most citrus species. Xanthomonas campestris pv. campestris (Xcc) is the causal agent of black rot disease in cruciferous plants. In Xcc, cell-cell signaling is mediated by diffusible signal factor (DSF). Synthesis of DSF depends on RpfB and RpfF. DSF perception and signal transduction have been suggested to involve a two-component system comprising RpfC and RpfG. It has been proposed that these proteins participate in a signal transduction system linking changes in the environment to the synthesis of DSF and the expression of virulence genes. Although the cluster of the rpf genes in Xac has synteny with the corresponding cluster in Xcc, two genes (rpfH and rpfI) are absent in Xac. To investigate DSF-mediated regulation during Xac-Citrus limon interaction, we constructed two strains of Xac, one with a mutation in the rpfF gene, leading to an inability to produce DSF, and one with a mutation in the rpfC gene leading to an overproduction of DSF. These mutants also show decreased levels of extracellular cyclic β-(1,2)-glucans and decreased production of endoglucanase and protease extracellular enzymes. The Xac DSF-deficient rpfF and the DSF-hyper producing rpfC mutants are both severely compromised in their ability to cause canker symptoms in lemon leaves compared to the wild-type. Here we provide evidence that rpf genes in Xac are involved in controlling virulence factors mediated by DSF. Keywords: canker, DSF, extracellular polysaccharide, quorum sensing. Abbreviations: DSF: diffusible signal factor Citrus canker is an endemic disease of citrus in Argentina. Invasion and colonization of the host mesophyll is accomplished through natural openings and plant tissue lesions (Graham et al. 2004). Most commercial citrus cultivars are moderately to highly susceptible to Xac. Thus, there is considerable need for a better understanding of the process of infection to pave the way for the development of biotechnology strategies that will support sustainable management of the disease. Xcc is one of Xanthomonas species most extensively studied at the genetic level. The complete genome sequences of Xac and Xcc, revealed that the two bacteria have more than 80% of their genes in common and that their chromosomal gene order is largely conserved (da Silva et al. 2002). The ability of Xcc to elicit disease depends on several factors, including the synthesis of extracellular plant cell wall-degrading enzymes and the extracellular polysaccharide (EPS) xanthan (Tang et al. 1991). Production of these pathogenesis factors is regulated by a cluster of genes (A-I) called rpf (for regulation of pathogenicity factors) (Slater et al. 2000). Only two of these genes (rpfH and rpfI) are absent in Xac (da Silva et al. 2002). The predicted protein, RpfH, is structurally related to the sensory input domain of RpfC (Slater et al. 2000). RpfI positively regulates the synthesis of proteases, endoglucanases and EPS in Xcc (Dow et al. 2000). The locus corresponding to rpfI in Xac is occupied by a truncated copy of an insertion sequence (da Silva et al. 2002). The absence of rpfH and rpfI in Xac may be consistent with the distinct modes of entry of the two bacteria, with Xcc accessing the leaf mainly via hydathodes at the leaf margin and subsequently colonizing the xylem vessels (Wallis et al. 1973) and Xac entering primarily through the stomates and colonizing the apoplasm of the fruit, leaf and stem tissue (Graham et al. 2004). The major aim of this work is to identify genes involved in Xac pathogenicity, in particular, genes mediating cell-cell signaling. Our primary approach has been the isolation of additional mutants affected in the synthesis and perception of DSF. Such studies may provide a rational basis for the development of crop protection methods based on interference with intercellular signaling events. Bacterial strains and culture conditions The wild-type (8004) and rpfF (8523) and rpfC (8557) mutant strains of Xcc were described previously (Tang et al. 1991; Slater et al. 2000). The wild-type strain of Xac was obtained from cankerous leaves of infected lemon trees in the province of Tucumán, Argentina. Xcc and Xac strains were cultured in peptone-yeast extract-malt extract (PYM) nutrient medium (Vojnov et al. 2001), supplemented with D-glucose at a final concentration of 2% (w/v) at 28ºC. For growth on plates, PYM was solidified with 1.5% (w/v) agar to make PYMA. L medium contained bactotryptone, 10 g/l; yeast extract, 5 g/l; sodium chloride, 5 g/l and D-glucose, 1 g/l. E. coli strains were grown at 37ºC in Luria-Bertani (LB) broth and on LB agar (Dow et al. 2003). Bacterial growth was measured in an spectrophotometer at 600 nm. When required, the antibiotics, Rifampicin (Rif) and Kanamycin (Km) were added to the growth media at concentrations of 10 µg/ml and 50 µg/ml, respectively. Site-directed gene replacement of the Xanthomonas axonopodis pv. citri Genomic DNA from the Xac strain was extracted according to Chen and Kuo (1993). PCR products, from the region encoding the wild-type Xac rpfF and rpfC genes, were amplified using primers designed from Xac sequences available in GenBank (da Silva et al. 2002) and cloned into the pGEM-Teasy vector (Promega). A cassette containing a Km resistance gene was inserted in the rpfF and rpfC cleavage sites, generating deletions in the rpfF and rpfC gene. These constructs were digested and cloned into the suicide vector pSac (Quandt and Hynes, 1993) and transformed into wild-type Xac by electroporation. Transformed bacteria were selected on PYM media supplemented with 5% sucrose and 50 µg/ml Km. Disruption of the rpfF and rpfC loci in the mutant strains was confirmed by PCR and Southern blot (data not shown). Bioassay of DSF extracted from culture supernatants DSF was extracted into ethyl acetate from culture supernatants of Xcc or Xac strains grown in PYM as described by Barber et al. (1997). The DSF bioassay is based on DSF's ability to restore endoglucanase production to Xcc rpfF mutants as described in Barber et al. (1997). Assays of extracellular enzymes and EPS For measurement of protease and endoglucanase activity, Xac strains were grown in PYM medium. Enzyme activity in cell-free culture supernatants were measured by radial diffusion assays into substrate-containing agar plates as described by Slater et al. (2000). For measurements of EPS production, strains were grown in PYM medium supplemented with 2% (w/v) D-glucose for 24 hrs. EPS was precipitated from culture supernatants by ethanol, dried and weighed as described by Tang et al. (1991). Methods to isolate the cyclic glucan from culture supernatants, for size-exclusion chromatography on BioGel P4, have been described previously (Vojnov et al. 2001). Bacterial cultures were washed and resuspended in water to a concentration of 108 cfu ml-1. Samples of these suspensions (50 µl) were spotted onto plates and allowed to dry before growth at 30ºC. Plates were stained when necessary as described in Tang et al. (1991). Plant preparation and measurement of bacterial growth in plant Citrus limon variety Eureka was grown in a greenhouse at a temperature of 18-25ºC. Xac and Xac mutants were grown in PYM media supplemented with the appropriate antibiotics at 28ºC for 24 hrs. Cells were resuspended to a concentration of 106 cfu ml-1. Xac strains were assayed for pathogenicity by inoculation of bacteria into the lamina of mature and young lemon leaves using three different procedures, namely pressure infiltrating with a 2 ml syringe without a needle, nicking the underside of the leaves with a razor blade followed by spraying, and spraying without nicking. The last method mimics the natural Xac infection process, in which bacteria enter to the leaf through openings in the leaf (via stomata), followed by colonization of the apoplasm. Inoculated plants were maintained for 35 days in a growth cabinet, with temperatures ranging from 25-28ºC, high humidity, a photoperiod of 16 hrs light, and a light intensity of 150 a 200 µE/sm2. Disease progression was monitored phenotypically and through analysis of bacterial growth curves (Siciliano et al. 2005). Isolation of Xanthomonas axonopodis pv. citri mutants with altered levels of DSF The mutant strains ΔrpfF and ΔrpfC of Xac were screened for protease and endoglucanase activities, as well as for levels of cyclic β-(1,2)- glucan and xanthan. The proteolytic activity was scored by detecting degradation of milk proteins, seen as a zone of clearing around the colonies. As shown in Figure 1a, the halo around the colony of the rpfF and rpfC mutants is reduced compared with the wild-type Xac strain. The activities of endoglucanase, an enzyme involved in plant cell wall degradation, was measured by a plate diffusion assay, where the zones of clearing indicate degradation of carboxymethylcellulose (CMC). Similar to protease activity, endoglucanase activity was found to be reduced in both rpfF and rpfC mutants (Figure 1b). As shown in Figure 1c and Figure 1d, cyclic β-(1,2)- glucan and xanthan in the XacΔrpfF and ΔrpfC mutants are reduced compared to wild-type Xac. Based on these results and given that these pathogenicity factors are induced by DSF, we can infer that DSF levels are altered in the Xac ΔrpfF and ΔrpfC mutants. Xac DSF reverses the phenotype of Xcc rpfF mutants Strains of Xcc carrying mutations in the rpfF and rpfC genes grew as matrix-enclosed aggregates in L medium, whereas the wild-type Xcc strain grew in a dispersed fashion. (Dow et al. 2003). We investigated the behavior of the Xcc ΔrpfF (8523) and ΔrpfC (8557) mutants in the presence of DSF extracts taken from the Xac ΔrpfC mutant. As shown in Figure 2a, wild-type Xcc grows in a dispersed fashion, but Xcc rpfF and rpfC mutants do not. Addition of DSF extracted from the Xac ΔrpfC mutant reverses the phenotype of mutant 8523, but not mutant 8557. Based on these findings, we conclude that the Xac ΔrpfC mutant produces DSF, which induces bacteria to grow in a dispersed manner, and that DSF from Xcc and Xac likely share structural similarities. To confirm that Xac is able to produce DSF, we analyzed DSF extracted from wild-type and Xac mutants using the DSF bioassay developed by Barber et al. (1997), which relies on the restoration of endoglucanase activity to an rpfF mutant strain of Xcc. Ethyl acetate extracts of culture supernatants from mutant and wild type strains were assayed for DSF activity. As expected the extracts of the Xac ΔrpfC mutants show bigger halos compared with the wild-type, while the rpfF strain extracts show no halos indicative of CMC degradation (Figure 2b). Extracts from Xcc DSF-hyper producing rpfC and DSF-non-producing rpfF mutant strains were used as controls (Figure 2b). Reduction of virulence on lemon leaves is preferentially associated with mutation of the rpfC gene in Xac To test whether the mutations in the rpfF and rpfC genes of Xac affect general fitness in plant, bacteria suspensions were inoculated at a concentration of 106 cfu ml-1, followed by measurements of bacterial growth and observation of canker symptoms. Figure 3 shows symptoms on lemon leaves 35 days post-infection (d.p.i.) using the methods described above (nick and spray, A; spray alone, B; and pressure infiltration, C) with Xac ΔrpfF and ΔrpfC mutants and wild-type Xac. Although both mutants show reduced pathogenicity compared with wild-type, the DSF-hyper producing mutant (rpfC) is more severely compromised in its ability to cause canker disease in citrus. These results are most clearly evident with the spray inoculation method (B). Plants inoculated with MgCl2 show no disease symptoms. Further study of DSF`role in the plant inmune response and other regulatory mechanisms should advance our understanding of the adaptation of bacteria to parasitic life within plants and may allow us to develop tools to control Xanthomonas infection that function across plant species. |
| |||||||||

{kind=link}
{kind=link}
{kind=link}