 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
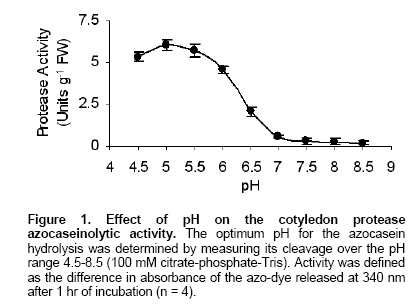
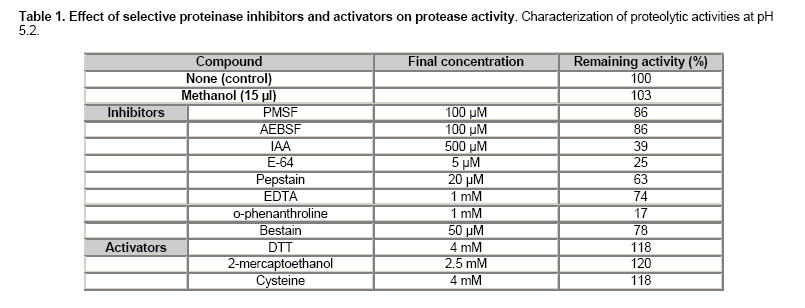
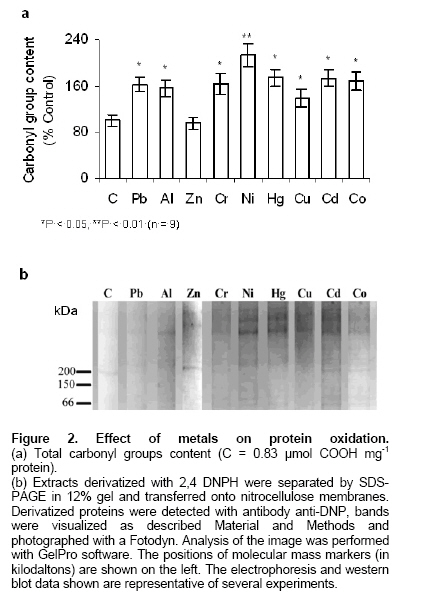
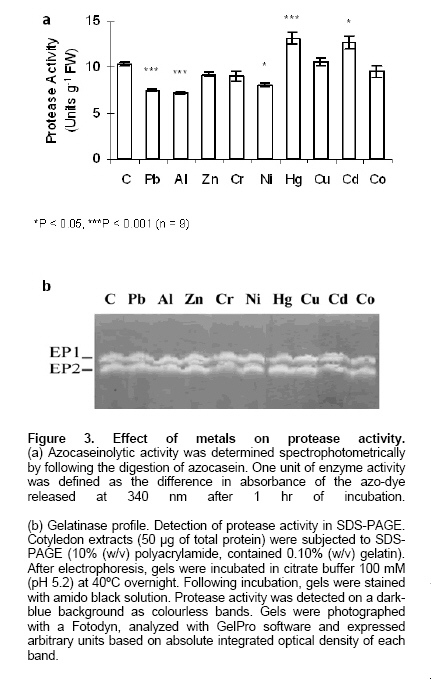
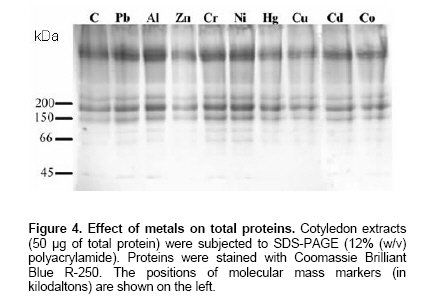
Electronic Journal of Biotechnology, Vol. 9, No. 3, June, 2006, Supp. 1, 2006, pg. 258- 262 RESEARCH ARTICLE Effect of different metals on protease activity in sunflower cotyledons Liliana B. Pena1 , María L. Tomaro2 , Susana M. Gallego*3 1Departamento de Química Biológica,
Facultad de Farmacia y Bioquímica,
Universidad de Buenos Aires,
Junín 956, (C1113AAC),
Buenos Aires, Argentina
Tel: 54 11 49648237
E-mail: lpena@ffyb.uba.ar Financial support: Grants from the Universidad de Buenos Aires (UBA) and from Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET) (Argentina). Code Number: ej06097 Abstract Proteases are crucial for living cells and play a role in plant cell adaptation to environmental conditions. Oxidative stress produced oxidized proteins which are selectively degraded by proteases. To understand the role of proteolysis in response to metal stress, sunflower plants (a plant suitable for phytoremediation) were treated with 100 µM of CdCl2, CuCl2, AlCl3, CoCl2, PbCl2, CrCl3, NiCl2, HgCl2 or ZnCl2. Changes in protease activity, gelatinase profile and protein oxidation were examined in sunflower cotyledons. Our results indicate that this tissue has mainly acid proteases belonging to different classes. Although all metals (except Zn) increased protein oxidation (62, 57, 112, 74, 74, 68, 64 and 40% for Pb, Al, Ni, Cd, Hg, Co, Cr and Cu over the control), they altered proteolysis in different ways. Pb, Al and Ni treatment decreased protease activity 22, 28 and 30% respect to control while Cd and Hg increased this activity in 23 and 27%. In Zn, Cu and Co treatments protease activity remained similar to control treatment. These results indicate that different proteases are involved in plant defence against metal toxicity. However, the identification of specific oxidized proteins involved in this process and the metal effect on class specific proteases should provide greater information. Keywords: Helianthus annuus L., oxidative stress, proteins degradation. Abbreviations:2,4 DNPH: 2,4-dinitrophenylhydrazine Plant genomes encode hundreds of proteases, but little is know about what roles they play in the life of a plant (Beers et al. 2004). Peptidases are classified based on their catalytic mechanisms. Five catalytic classes (serine, cysteine, aspartic, threonine and metallopeptidases) can been identified within the Arabidopsis genome sequence (Rawling et al. 2004). In general, proteases are thought to be involved in a range of processes, including germination, senescence, defence responses and programmed cell death (Fontanini and Jones, 2002; Segarra et al. 2002; Roberts et al. 2003; Coffeen and Wolpert, 2004). In addition, proteases might also play a role in the plant adaptation to changing environmental conditions. Proteolysis is also associated to oxidative stress promoted by ROS (O2-, H2O2, OH). The protein modification exerted by oxidative stress is characterized for the production of carbonyl groups in the molecules (Palma et al. 2002). Sunflower is a plant that dramatically reduces levels of metals in the soil, so it is suitable for phytoremediation (Salt et al. 1995). To understand the role of proteolysis in response to metal stress, sunflower plants were treated with 100 µM of different metals. Changes in proteolytic activity, gelatinase profile and protein oxidation have been examined in sunflower cotyledons, a tissue where proteolysis is essential to mobilize protein reserves and thus supplying aminoacids for protein synthesis in developing tissues. Plant material and growing conditions Seeds of Helianthus annuus L. (cv Paraiso 20) were germinated in vermiculite and grown in a controlled climate room (24 ± 2ºC, 50% relative humidity, photoperiod of 16 hrs, light intensity 300 µmol m-2 s-1). Ten days seedlings were removed from the pots, roots were carefully washed and transferred to separated containers (3 L) for hydroponics, 20 plants per container. The hydroponics medium was half strength Hoagland's nutrient solution (Hoagland and Arnon, 1953) (control) or containing 100 µM of CdCl2, CuCl2, AlCl3, CoCl2, PbCl2, CrCl3, NiCl2, HgCl2, ZnCl2, (pH 5.8) continuously aerated. After 4 days of treatment cotyledons were used for all further determinations. Extracts were prepared from 1 g of cotyledons homogenized in 5 ml of 10 mM buffer phosphate-K (pH 7.5), 2 mM cys, 5 mM 2-mercaptoethanol, 0.1 g PVP and 0.1% (v/v) Triton X-100 at 4ºC. The homogenates were centrifuged at 10000 g for 20 min and the supernatant fraction was used for the assays. Protease activity was measured using azocasein as substrate (Weckenmann and Martin, 1984). Activity was defined as the difference in absorbance of the azo-dye released at 340 nm after 1 hr of incubation. The optimum pH for the azocasein hydrolysis was determined by measuring its cleavage over the pH range 4.5-8.5 (100 mM citrate-phosphate-Tris). To test the effect of inhibitory or activating compounds, control extracts were preincubated for 30 min at 40ºC in the presence of each metals, or in the presence of the following compounds: PMSF, AEBSF, IAA, E-64, EDTA, o-phenanthroline, pepstain, bestatin, DTT, 2-mercaptothanol, or cys, after which the substrate was added to start the reaction. To measure protease activity SDS-gelatin-PAGE was used. Regular SDS-PAGE (Laemmli, 1970) was performed, except that the 10% resolving gel contained 0.10% (w/v) gelatin. Aliquots containing equal protein amount (50 µg) were mixed with non-reducing sample buffer (0.1 M Tris-HCl, pH 6.8, 2% (w/v) SDS, 10% (v/v) glycerol, 0.01% (w/v) bromophenol blue). Following electrophoresis at 4ºC, gels were transferred to 1.5% (v/v) Triton X-100 in aqueous solution for 1 hr at 20ºC to allow removal of SDS and renaturation of proteases. Gels were incubated in citrate buffer 100 mM (pH 5.2), 1 mM CaCl2 2 h at 40ºC. Then gels were stained with 0.1% (w/v) amido black, 40% (v/v) methanol, 10% (v/v) acetic acid for 2 hrs and distained with 40% (v/v) methanol, 10% (v/v) acetic acid. Protease activity was detected on a dark-blue background as colourless bands. Proteins and carbonyl groups content Protein concentration was determined according to Bradford (1976); bovine serum albumin was used as standard. SDS-PAGE was performed using 12% (w/v) gel. Proteins were visualized by Coomasie Brilliant Blue R-250-staining. Protein oxidation was measured as total carbonyl groups content by their reaction with 2,4 DNPH (Levine et al. 1990). Extracts derivatized with 2,4 DNPH were separated by SDS-PAGE in 12% (w/v) gel and transferred onto nitrocellulose membranes. Derivatized proteins were detected with antibody anti-DNP. Bands corresponding to oxidized proteins were visualized using secondary antibodies conjugated with HRP and DAB as substrate. Values are mean values ± s.e. Differences among treatments were analyzed by 1-way ANOVA, taking P < 0.05 as significant according to Tukey's multiple range test. Protease exhibited high azocaseinolytic activity at acid pH (maximum between 5.0-5.5) (Figure 1), and optimal temperature occurred between 35 and 45ºC (data no shown). The characterization of proteolytic activities at pH 5.2 with class-specific inhibitors revealed that proteases belonging to cysteine, aspartate and metalloprotease groups were detected. These inhibitor studies indicate the involvement of various proteases in cotyledon protein metabolism. Sulphydryl groups protectors activated protease activity (Table 1). Protein carbonylation is an irreversible oxidative process leading to a loss of function of the modified proteins. These oxidized proteins are selectively recognized and degraded by proteolytic enzymes (Palma et al. 2002). Extreme environmental conditions that induce oxidative stress have been associated to an increased carbonyl groups content and to an induction in protease activity. In our experimental condition, although metals produced oxidative stress (except Zn) and increased protein carbonylation (Figure 2a), they had different effect on total protease activity. While Pb, Al and Ni decreased protease activity (22%, 28% and 30% respectively, respect to control values), Cd and Hg increased it (23% and 27% respectively, over the control values) (Figure 3a). Co and Cu increased carbonyl groups content (Figure 2a) (68% and 40% over the control), however protease activity remained as in control treatments. Some metalloproteinases contain Zn, Co or Mn to activate the water molecule for the hydrolysis of peptide bonds, that is probably why Zn and Co did not cause modification in protease activity. Metals and auto digestion process themselves had not effect on protease activity (data not shown). Imposition of metals on sunflower plants, although altered pre-existing proteases activities (EP1 and EP2), did not induce novel proteases in cotyledons (Figure 3b). Immunodetection of carbonyl residues showed increased oxidation of protein with high molecule weight (Figure 2b). Metal treatments had not significant effect on protein content and profile (Figure 4). In conclusion, sunflower cotyledons have mainly acid protease activity belonging to different classes of proteolytic activities. Degradation of oxidized proteins removes potential toxic fragments and provides aminoacids for new protein synthesis. Metals may affect plant metabolism in different ways. Pb, Al and Ni toxicity mechanism may be attributed to inhibition of protease activity, this seems to be causing carbonylation of proteins. On the other hand, Cd and Hg induce oxidative stress and enhance proteolytic activity. Due to Zn and Co are cofactors of metalloproteases, they have no effect on its activity. These results indicate that different proteases are involved in plant defence against metal toxicity. More studies on the identification of the specific oxidized proteins and the effect of metals on class specific proteases are necessary to improve our knowledge of the mechanisms involved in the different metals toxicity and the possible use of proteases to generate metal tolerant transgenic plants |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}