 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Electronic Journal of Biotechnology, Vol. 9, No. 3, June, 2006, Supp. 1, 2006, pg. 267-271 RESEARCH ARTICLE Identification of microsatellites linked to Lr47 Leonardo Sebastián Vanzetti1 , Juan Carlos Brevis2 , Jorge Dubcovsky3 , Marcelo Helguera*4 1Grupo de Biotecnología y Rec. Genéticos,
INTA EEA Marcos Juárez,
Ruta 12 S/N, (2580) Marcos Juárez,
Córdoba, Argentina
Tel/Fax: 54 3472 425001
E-mail: lvanzetti@mjuarez.inta.gov.ar Financial support: Grants from the Government of Argentina, Proyecto Nacional INTA 52-520209 and in the US by competitive grant IFAFS 2001-04462. Code Number: ej06099 Abstract Leaf rust resistance gene Lr47 is located within a interstitial segment of Triticum speltoides Taush. 7S chromosome translocated to the short arm of chromosome 7A of bread wheat. This gene is resistant against currently predominant races of leaf rust from Argentina. The objectives of this study were to identify microsatellites linked to this source of resistance as a potential tool to introgress this source of resistance. Isogenic lines with and without Lr47 developed from 10 cultivars/breeding lines as well as 10 microsatellites previously mapped in 7AS chromosome were used in this study. Microsatellite gwm 60 was the only marker that co-segregated completely linked to Lr47. These data indicate that gwm 60 could be a valuable marker to introgress Lr47 in wheat germplasm. Keywords: gwm 60, leaf rust, marker assisted selection, Puccinia triticina Ericks., Triticum aestivum.
Leaf rust (caused by Puccinia triticina Ericks.) is one of the mayor diseases of wheat worldwide. In South America, yield losses due to leaf rust occur annually throughout the hard red spring wheat regions of Argentina, Chile, Uruguay, Brazil and Paraguay. Incorporating genetic resistance to this pathogen into adapted germplasm is a mayor goal in wheat breeding programs, as using disease resistance genes minimizes the need for the application of costly fungicides, thus reducing environmental contamination risks and decreasing production costs. Unfortunately, the gene pool of cultivated wheat for resistance to pests and pathogens is inadequate to respond to the evolution of different pathogen populations. Replacement of highly variable land races by a short number of high yielding, pure-line varieties in many parts of the world, including the South Cone, has reduced the wheat gene pool. In this context, genes from wild wheat relatives have contributed to increase the genetic diversity of hexaploid wheat in genes conferring resistance against leaf rust (Puccinia recondita Rob. ex Desm.), stem rust (Puccinia graminis Pers.), Hessian fly [Mayetiola destructor (Say)], Russian wheat aphid (Diuraphis noxia Mordvilko) between other pathogens (reviewed by McIntosh, 1991). At least 51 leaf rust resistance genes from wheat and wheat relatives have been catalogued (McIntosh et al. 2003) and molecular markers are available for many of them (Robert et al. 1999; Helguera et al. 2000; Helguera et al. 2003; Helguera et al. 2005; http://maswheat.ucdavis.edu/, verified August 20, 2005). In some cases, resistance genes derived from wild relative translocations are not deployed in breeding programs because the linkage between targeted genes and undesirable genes on the alien segment usually result in yield and/or quality penalties. A good example is the Lr19 translocation including the undesirable linked yellow pigment gene (Prins et al. 2001; Zhang et al. 2005). Consequently, to be deployed in agriculture the alien chromosome segments should be as short as possible. The identification of alternative markers mapping in wild relatives translocations is a useful tool to generate shorter translocations carrying the desirable character without linked undesirable traits. Triticum speltoides Taush (2n=14, S genome) is an attractive source of high levels of resistance to leaf, stem and stripe rust of wheat (Dvorak, 1977), and leaf rust resistance genes Lr28, Lr35, Lr36, Lr47 and Lr51 were derived from this species (McIntosh et al. 2003; Helguera et al. 2005), as different T. speltoides chromosomes translocated to different wheat chromosomes. The leaf rust resistance gene Lr47 is located within a segment of Triticum speltoides Taush. chromosome 7S transferred to the chromosome 7A of hexaploid wheat into a interstitial translocation 20-30 cM long (Dubcovsky et al. 1998). The 7AS-7S#1S-7AS.7AL T. speltoides translocation was characterized using 14 RFLP markers previously mapped on the short arm of chromosome 7Am of T. monococcum (Dubcovsky et al. 1996) finding 7 RFLP loci that mapped into the translocation (Dubcovsky et al. 1998). RFLP markers are expensive and time consuming, therefore PCR based markers are an attractive alternative for breeding programs. Unfortunately, only PCR markers derived from the RFLP ABC465 (maps close to the last part of the 7AS-7S#1S-7AS.7AL translocation, close to the centromere) are available (Helguera et al. 2000). An interesting source of PCR markers in wheat are the microsatellites (Tautz and Renz, 1984). These loci are amplified using primers (18-25 bp long) specific for sequences flanking hyper variable regions of tandem repeats of 2-4 base pairs. Several molecular maps including more than 320 microsatellites covering the whole wheat genome are available (Bryan et al. 1997; Roder et al. 1998; Paillard et al. 2003). The aim of the present work was to characterize the 7AS-7S#1S-7AS.7AL T. speltoides translocation using microsatellites previously mapped on 7A wheat chromosome to identify novel PCR markers linked to Lr47 . |
One or more isogenic lines homozygous for Lr47 developed from 9 wheat cultivars and/or breeding lines from Argentina and USA were used in this study. The 7AS-7S#1S-7AS.7AL T. speltoides translocation carrying Lr47 was introgressed in Prointa Puntal (hard red winter from Argentina), Prointa Oasis and Prointa Imperial (hard red springs from Argentina), Kern, Yecora Rojo and Express (hard red springs from USA), UC1037, UC1041 and RS15 (University of California breeding lines from USA), by marker assisted backcrossing (BC). Each cultivar/breeding line was crossed with Pavon-Lr47 (donor of the 7AS-7S#1S-7AS.7AL T. speltoides translocation) and the nine F1s were backcrossed with the respective parents. In each of the 4 BC generations (or 6 in Kern, Yecora Rojo, Express, UC1037, UC1041 and RS15) two individuals heterozygous for the 7AS-7S#1S-7AS.7AL translocation were selected by marker assisted selection using Lr47 specific markers according to Helguera et al. (2000). Finally, BC4 (or BC6) plants heterozygous for 7AS-7S#1S-7AS.7AL translocation were self-pollinated and homozygous 7AS-7S#1S-7AS.7AL plants were selected from BC4 F2 (or BC6 F2) using codominat Lr47 markers (Helguera et al. 2000). These selected BC4 F2 plants are expected to be 96.8% (or more than 99% in BC6 F2) identical to the recurrent parent and homozygous for the Lr47 gene. The nine recurrent parents, the breeding line Pavon-Lr47, Pavon without the 7AS-7S#1S-7AS.7AL translocation (hard red winter from CIMMYT) and Chinese Spring nullisomic - tetrasomic lines N7A N7B and N7D (Sears, 1954) were also used in this study. A diverse set of varieties from Argentina including Biointa 3001, Buck Pingo, ACA 302, Prointa Bon. Hurón, Klein Don Enrique, Baguette 10, Relmó Tijereta, ACA 223, Klein Estrella, Buck Biguá, Buck Mataco, Prointa Cinco Cerros and Klein Volcán was analyzed to validate gwm 60 as a marker for Lr47.
Tests for resistance to leaf rust
Isolines carrying Lr47 from Prointa Puntal, Prointa Oasis and Prointa Imperial were grown in Marcos Juárez (Argentina) as part of the National Institute of Agricultural Technology (INTA) Wheat Breeding Program during years 2000, 2001, 2002 and 2003 to evaluate leaf rust resistance against local races of the pathogen. In the field experiment, recurrent parents (highly susceptible against local leaf rust races) were also included as controls.
DNA extraction and PCR markers
Genomic DNA was extracted from fresh leaves of single plants using a fast, small-scale DNA isolation procedure based on Wining and Langridge (1991).
A total of 10 wheat microsatellites previously mapped on 7A chromosome were included in this study: gwm 60, gwm 635 and wms 666 from Röder et al. (1998); wmc 168, wmc 497, wmc 83 and wmc 17 from the Wheat Microsatellite Consortium (Gupta et al. 2002); Barc 70, Barc 174, and Barc 154 (Somers et al. 2004).
The PCR reactions were performed in 25 µl aliquots in a PTC-100 (MJ Research) thermal cycler. The reaction buffer contained 100 ng of template DNA, 1X Taq polymerase buffer (Promega Corp. Madison, WI), 1.0 U Taq (Promega), 0.2 mM of each deoxynucleotide, 0.2 µM of each primer and 1.5 mM of MgCl2. Forty cycles of 45 sec at 94ºC, 45 sec between 55 to 65ºC depending on the primer combination and 1 min at 72ºC were performed, followed by an elongation step of 10 min at 72ºC. PCR reactions (10 µl each) were run on 8% polyacrylamide gels 15 cm long (Ruby SE600, Amersham BioSciences, CA), stained with ethidium bromide [0.5 g/L] and visualized with UV.
Infection types to Puccinia triticina
Field tests carried out in Marcos Juárez (Argentina) during years 2002, 2003 and 2004, showed that the presence of Lr47 in BC2 F2 isolines from cvs. Prointa Puntal, Prointa Oasis and Prointa Imperial improved the resistant to local leaf rust isolates relative to the recurrent parents. In all tested years, isolines carrying Lr47 showed resistance against local leaf rust races while recurrent parents showed moderately susceptible or susceptible responses. The host responses observed in isolines carrying Lr47 agree with previous data obtained from Pavon-Lr47 showing resistance against nine leaf rust races (Dubcovsky et al. 1998). This novel source of leaf rust resistance has not been used in commercial wheat cultivars from South America. The good performance of Lr47 against local strains of leaf rust isolates suggests that this gene would be a useful source of resistance against this pathogen in wheat.
Molecular characterization of the 7AS-7S#1S-7AS.7AL T. speltoides translocation using microsatellites
Based on electrophoretic conditions used in this study, four of the ten tested microsatellites (gwm 635, wmc 17, wms 666, wmc 83) were monomorphic, considering Pavon, Pavon-Lr47 and the isolines with/without Lr47 from the 9 cultivars/breeding lines used in this study, so no additional information was obtained from them.
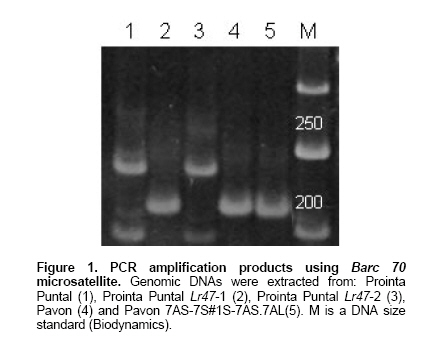
Another group of microsatellites (Barc 70, wmc 168, Barc 174, wmc 497, Barc 154), was monomorphic for Pavon and Pavon-Lr47 but polymorphic when isolines with and without Lr47 were considered in the analysis, suggesting that this group of markers have a physical position outside the 7AS-7S#1S-7AS.7AL translocation. This hypothesis could be corroborated in microsatellite Barc 70. Using this marker, Pavon and Pavon-Lr47 amplified a fragment of approximately 220 bp (lanes 4 and 5 in Figure 1), the cultivar Prointa Puntal showed a approximately 240 bp fragment (lane 1) and isolines Prointa Puntal Lr47-1 and Prointa Puntal Lr47-2 showed fragments of 220 and 240 bp (lanes 2 and 3, Figure 1).
The most probable explanation of this result is the occurrence of a crossing over between Lr47 and Barc 70 in Prointa Puntal Lr47-2. This can be possible only if Barc 70 is outside of the 7AS-7S#1S-7AS.7AL translocation, as the recombination between 7A and 7S chromosomes is prevented in presence of the Ph1 gene (Helguera et al. 2000). Similar data was obtained from microsatellites wmc 168, Barc 174, wmc 497, Barc 154.
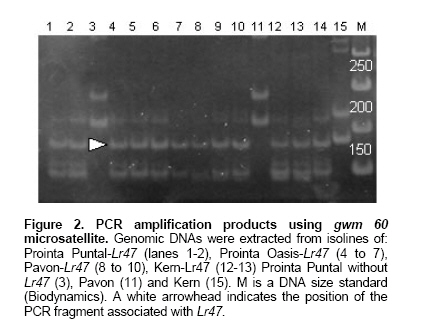
The only marker that showed clear polymorphisms between Pavon and Pavon-Lr47 was gwm 60 (lane 11 and lanes 8-10, respectively in Figure 2), suggesting that it maps into the 7AS-7S#1S-7AS.7AL translocation. Nulli-tetrasomic analysis confirmed that the polymorphic fragment amplified by gwm 60 belongs to 7A chromosome (data not shown). When gwm 60 was evaluated in isolines with and without Lr47, a fragment of approximately 180 bp was observed only in isolines carrying Lr47 (Figure 2). In isolines without Lr47 PCR fragments of size different from 180 bp were amplified in all cases (lanes 3, 11 and 15 in Figure 2).
The scoring obtained with gwm 60 considering at least 30 isolines with and without Lr47 used in this study is identical to the scoring obtained using Lr47 markers developed by Helguera et al. (2000) and used in the backcrossing program that generated the isolines, this data demonstrates that gwm 60 is a good alternative marker to introgress Lr47 and a useful tool for the future dissection of this T. speltoides chromosome segment. To validate this marker in a larger set of wheat cultivars, 13 wheats from Argentina were evaluated (see Plant Materials section) and none of the tested cultivars amplified the 180 bp fragment associated with 7AS-7S#1S-7AS.7AL translocation.
The Xgwm60 locus has been mapped in chromosome 7A between loci Xabc158 and Xcdo475 (Röder et al. 1998), and Xcdo475 is at least 16-cM away from Xabc465 (Dubcovsky et al. 1998), which is the RFLP locus previously converted to a PCR marker for Lr47 (Helguera et al. 2000). The fact that loci Xgwm60 and Xabc465 are not so closely linked would be helpful to select shorter 7S chromosome segments carrying Lr47 through a second round of homologous recombination. Preliminary results suggest that the T. speltoides chromosome segment has a negative effect on flour yield, and therefore it will be desirable to separate this negative effect from Lr47. The gwm 60 marker reported here will be a useful tool to achieve this objective.
The authors acknowledges to Gabriela Tranquilli and Carlos Bainotti for their valuable suggestions in the writing of the manuscript.
Note: Electronic Journal of Biotechnology is not responsible if on-line references cited on manuscripts are not available any more after the date of publication.
Supported by UNESCO / MIRCEN network
© 2006 by Pontificia Universidad Católica de Valparaíso -- Chile
| |||||||||

{kind=link}
{kind=link}