 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Electronic Journal of Biotechnology, Vol. 9, No. 3, June, 2006, Supp. 1, 2006, pg. 276-280 RESEARCH ARTICLE Meiotic study of Zea mays ssp. mays (2n = 40) x Tripsacum dactyloides (2 n = 72) hybrid and its progeny María del Carmen Molina*1 , María Dina García2 , Silvia Elena Chorzempa3 1Instituto Fitotécnico de Santa Catalina,
Facultad de Ciencias Agrarias y Forestales,
Universidad Nacional de la Plata,
CIGen (UNLP-CONICET-CIC),
Garibaldi 3400, CC4, 1836 Llavallol,
Buenos Aires, Argentina
Tel/fax: 54 11 4282 0233
E-mail: mcmgen@yahoo.com Financial support: FCAyF-UNLP (grant A129), FCA-UNLZ and CONICET (grant 4650). Code Number: ej06101 Abstract Maize (2n = 40) x Tripsacum dactyloides (2n = 72) F1 hybrid plants (2n = 56) were obtained by embryo rescue and induction of somatic embryogenesis/organogenesis. Hybrid plants showed Tripsacum-like phenotypes, tolerance to stresses such as NaCl salinity and low temperatures. The more frequent meiotic configurations were 28II (24%), 24II + 2IV (19%) and 26II + 1IV (12%), with an average per cell of 0.55I + 25.18II + 1.19IV. Significant differences between plants were not observed. Pollen fertility ranged from 0% to 50%. After pollination with maize or Tripsacum, 20% of F1 plants have developed viable seeds, which originated the progeny. Thirty five percent of the progeny showed 2n = 56 chromosomes and F1 like-phenotypes, which suggests they have apomictic origin. The remaining plants were fertile and they showed maize-like phenotypes and different chromosome numbers (2n = 22, 24, 26, 28 and 30), because they kept the complete maize chromosome complement and some of the Tripsacum chromosomes. Meiotic cells showed pairing between chromosomes from both parental species, which suggests the possibility of genetic recombination between them. Keywords: chromosome number, corn, eastern gamagrass, remote hybridisation.
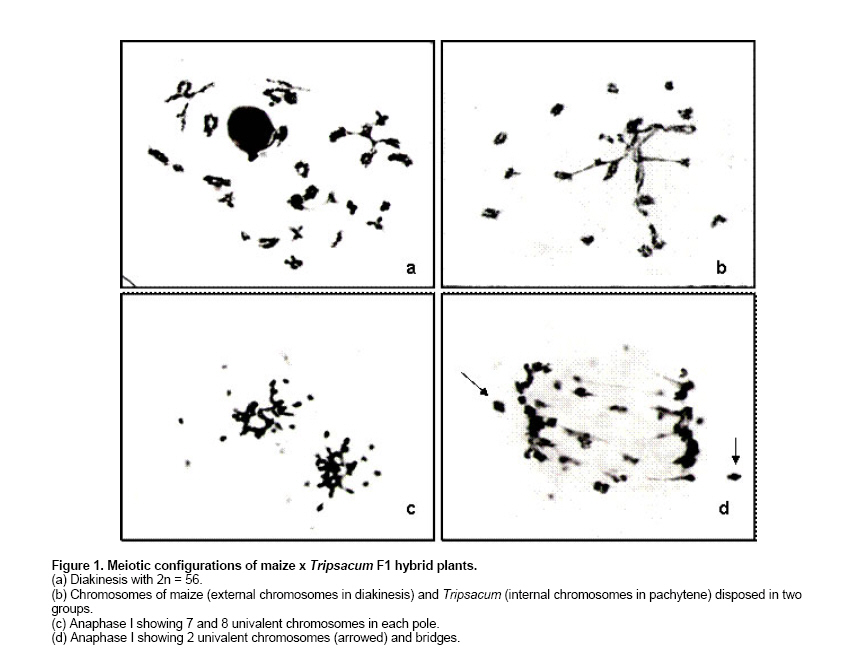
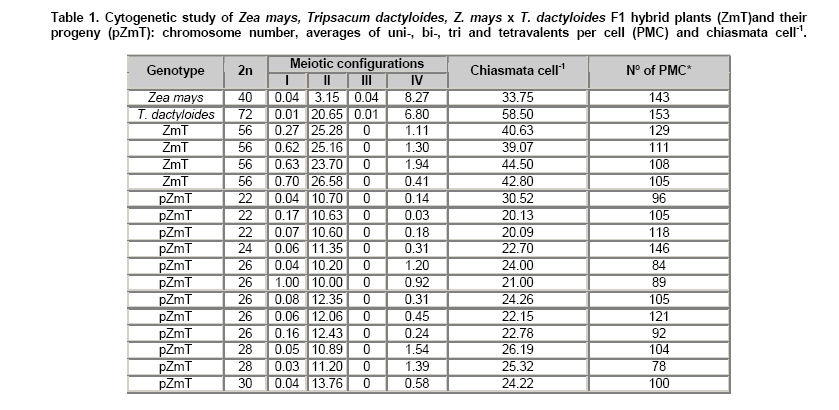
Interspecific hybridisation of remotely related plant species offers a great potential to improve efficiency in crop breeding and production. Introduction of novel genetic variability and apomictic seed development into crops have been recognizable as two important achievements of the manipulation of reproductive systems in plants (Matzk et al. 1997). Corn (Zea mays ssp. mays) is an annual crop that shows susceptibility to adverse environmental conditions such as low temperatures (Hodges et al. 1995) and NaCl salinity (Pasternak et al. 1995). Tripsacum dactyloides is a perennial forage which presents high palatability and productivity (Faix et al. 1980), tolerance to different environmental stresses (Foy, 1997; Clark et al. 1998; Ray et al. 1998) and apomictic reproduction (Burson et al. 1990; Leblanc et al. 1995a; Kindiger et al. 1996a; Grimanelli et al. 1998). Such characteristics make this species an interesting source of genetic variability for corn improvement. Different authors have investigated the possibility to introgress some characteristics from Tripsacum to maize, for example, disease tolerance (Bergquist, 1981), constitutive aerenchyma (Ray et al. 1999), and gametophytic apomixis (Leblanc et al. 1995b; Kindiger et al. 1996b; Lukina and Chistyakova, 1999). Hybrids between maize and Tripsacum can be obtained with more or less success, according to the maize genotype used as female parent, through embryo rescue and induction of organogenesis or somatic embryogenesis (Furini and Jewell, 1995; García and Molina, 1997). Maize x T. dactyloides F1 hybrid plants are sexually sterile, but low percentages of viable seeds have been obtained mainly by apomixis (Leblanc et al. 1995a; Sokolov et al. 2000). Regenerated hybrid plants showed salinity tolerance (Pesqueira et al. 2003). Further, F1 hybrid plants grown under field conditions or culture chambers showed high tolerance to low temperatures (-2ºC and 5ºC, respectively), although maize and Tripsacum showed great susceptibility under the same conditions (Jatimliansky et al. 2004). The objectives of this work were to evaluate pollen fertility and production of viable seeds of Z. mays ssp. mays (2n = 40) x T. dactyloides (2n = 72) regenerated hybrid plants, and to analyse meiosis of fertile F1 hybrid plants and the progeny of these plants. Inbred plants of Z. mays ssp. mays N107B (2n = 40, supplied by Maize Genetics Coop. Stock Center, Urbana, Illinois) were pollinated with Tripsacum dactyloides (2n = 72). Immature embryos originated embryogenic and organogenic callus cultures on the basic medium (García et al. 1992) supplemented with 4 μmolL-1 2,4 dichlorophenoxyacetic acid. Regenerated shoots were rooted on the basic medium free of plant growth regulators. The 130 F1 plants analysed in this work arose from one embryo. The progeny was obtained from free pollination of F1 plants with maize or Tripsacum. Immature tassels were fixed in Farmer solution (3:1 absolute ethanol-acetic acid) and coloured with a 2% solution of acetic haematoxylin. Pairing configurations were observed at diakinesis-metaphase I. Pollen fertility was determined using Lugol solution. Pollen was counted as fertile if at least 75% of the grain was coloured. Z. mays ssp. mays (2n = 40) x T. dactyloides (2n = 72) F1 hybrid plants showed a chromosome number 2n = 56, Tripsacum-like phenotypes, an average height of 2.10 m, 3 to 20 tillers per plant and 4 or 5 ears per stem. Their flowering period lasted from November to June or July. Half of the plants showed an annual cycle and the rest were biannual or perennial ones. Further, tolerance to low temperatures and NaCl salinity was observed in these plants (Pesqueira et al. 2003; Jatimliansky et al. 2004). Pollen fertility ranged from 0% (most of the plants) to 50%. Eighty percent of the plants did not produce viable seeds; the other 20% developed 2 to 10 viable seeds per ear. Normal microspore development but complete male sterility was observed in 56-chromosome maize x Tripsacum backcross hybrids (Kindiger, 1993). Over 40 plants were evaluated from the progeny, 35% showed 2n = 56 chromosomes (Figure 1a) and F1-like phenotypes. Pollen fertility ranged from 0% to 50%. The other 65% of the plants resembled maize more than Tripsacum phenotype, except that they showed up to 3 tillers per plant and continuous flowering on very prolific stems during the growing season. Pollen fertility ranged from 40 to 96%. Tetraploid (2n = 4x = 72) and triploid (2n = 3x = 54) eastern gamagrass reproduced by diplosporous pseudogamy, Antennaria-type megagametogenesis and diploid parthenogenesis (Burson et al. 1990). Chromosome number of 2n = 56 observed in 35% of the maize/Tripsacum F1 progeny suggests that these plants were originated by apomixis as described in poliploid Tripsacum species (Burson et al. 1990). However, reduction of Tripsacum chromosome complement was observed in 65% of maize/Tripsacum F1 progeny plants. A previous report (Kindiger et al. 1996b) also showed partial loss of Tripsacum chromosomes in backcrosses of an apomictic maize/Tripsacum F1 hybrid (2n = 56). The most frequent meiotic configurations in diakinesis - metaphase I (% of analysed cells), average number of uni-, bi-, tri-, and tetravalens and chiamata cell-1 (Table 1) were the following: Zea mays ssp. mays (2n = 40) showed 10IV (30%) and 9IV + 2II (24%), an average number of 8.15IV + 3.27II and 33.75 chiasmata cell-1. Cells showed normal anaphase I, i.e., a regular migration of 20 chromosomes to each pole. T. dactyloides (2n = 72) showed 5IV + 26II (21%) and 10IV + 16II (19%), an average number of 6.80IV + 20.65II and 58 chiasmata cell-1. F1 hybrid (2n = 56) showed 28II (24%), 24II + 2IV (19%) and 26II +1IV (12%), an average number of 0.55I + 25.18II + 1.19IV and 42 chiasmata cell-1. No significant differences were observed among analysed plants. Cells in anaphase I showed multipolar spindle, lagging chromosomes (Figure 1c) and one or two bridges (Figure 1d). Other meiotic abnormalities were also observed in F1 plants, such as 2 or more nuclei in different stages in the same cytoplasm, spatial separation and asynchronous stages of Tripsacum and maize chromosomes (Figure 1b). Some of the hybrid plants showed one or two tetravalents, one of them constituted by two chromosomes from maize and two from Tripsacum and the other (if there were two) by homoeologous Tripsacum chromosomes. F1 hybrid progeny plants with 2n = 56 and F1-like phenotypes showed cytogenetic characteristics similar to that described in F1 plants. Instead, the F1 progeny maize-like plants showed different chromosome numbers (2n = 22, 24, 26, 28 and 30) and produced viable seeds. The different chromosome numbers were the result of 26 to 34 Tripsacum chromosome elimination in metaphase I. Meiosis was normal in cells with 22 or 24 chromosomes. On the contrary, cells with 26 or more chromosomes showed lagging chromosomes, spatial separation of chromosomes in two groups by species and subsequent lost of Tripsacum chromosomes. The number of Tripsacum chromosomes represented as univalents was small in all MI cells of reduced maize-Tripsacum hybrids with 29 chromosomes (4.63 ± 0.17 to 6.72 ± 0.3 univalents per cell) (Lukina and Chistyakova, 1999) but higher than the number of univalents per cell observed in the present work for mazie x Tripsacum F1 hybrid (2n = 56) plants and their progeny (0.01 to 1.0 univalents per cell, Table 1). In conclusion, the study of maize x T. dactyloides F1 hybrid plants and their progeny showed that: i) Hybrids between different genera may express phenotypic characteristics absent in their progenitor species; ii) Fertility can be restored after most of the Tripsacum chromosomes elimination, iii) Pairing between parental species suggest the possibility of genetic recombination between them.
Note: Electronic Journal of Biotechnology is not responsible if on-line references cited on manuscripts are not available any more after the date of publication. © 2006 by Pontificia Universidad Católica de Valparaíso -- Chile The following images related to this document are available:Photo images[ej06101t1.jpg] [ej06101f1.jpg] |
| |||||||||

{kind=link}
{kind=link}