 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Electronic Journal of Biotechnology, Vol. 9, No. 3, June, 2006, Supp. 1, 2006, pg. 315-319 RESEARCH ARTICLE Rooting in Km selective media as efficient in vitro selection method for sunflower genetic transformation Laura Mabel Radonic1 , Julián Marcos Zimmermann2 , Diego Zavallo3 , Nilda López4 , Marisa López Bilbao*5 1Instituto de Biotecnología,
INTA Castelar ,
De los Reseros y Las Cabañas s/n (1712), Castelar,
Buenos Aires, Argentina,
E-mail: lradonic@cicv.inta.gov.ar| Financial support: This project was supported by the Agencia Nacional de Promoción Científica y Tecnológica (PICTO Nº 08-13164 and 08-12925), Argentina. Code Number: ej06108 Abstract Despite of numerous publications in sunflower genetic transformation, there is no efficient or reproducible protocol with low number of escapes. The latter would indicate that the selection method is not effective. In this work we used Km as selective agent, Agrobacterium tumefaciens EHA105 strain and a vector with the nptII gene under the nos promoter and uidA gene under 35S promoter. The response of agroinfected (A) and control (C) explants during the in vitro culture was studied and in both cases in presence or absence of Km in order to assign a differential morphologic response between transformed and non-transformed plants. The characteristics analyzed were: height, colour/aspect of the plantlets, in vitro rooting and in vitro bud-flower development. Selection was applied from the third regeneration media. Among the A plantlets two were capable of rooting, being positive by PCR, whereas the C were unable to root in presence of Km. One of them gave 6 seeds and in these plants, it was determined the presence of the transgene by PCR and GUS staining. This work shows that in Km selection, colour/aspect of shoots is not useful as selection criteria whereas rooting is an effective selection method in which no escapes were obtained. Keywords: genetic transformation, kanamycin, selection, sunflower. Transformation efficiencies have very low values, between 0.01 and 5%, so during the regeneration of transgenic plants, it is very important to set conditions where non-transformed cells or tissues are unable to grow and/or develop. This is even more important in the case of sunflower because in vitro regeneration of plantlets (T0) is via organogenesis, producing chimeras that not necessarily will give an offspring (T1) that carries and/or express the transgene, making the PCR analysis of the T0 not conclusive. For this reason, a selection gene is introduced along with the gene of interest, conferring the transformed cell the possibility to survive and regenerate in the presence of the selective agent (antibiotic or herbicide) whereas the non-transformed cells die or are unable to grow. Thus, the proper selection of a selective gene determines the possibility of success in the production of transgenic plants. Despite several publications in sunflower genetic transformation (Hewezi et al. 2002; Weber et al. 2003), there is no efficient or reproducible protocol with a low number of escapes, indicating that the selection method using Km is not effective. The aim of this work was to establish the use of the antibiotic Km as an efficient selective agent, trying to relate transgene presence with the differential in vitro behaviour.
Seeds from the public genotype HA89 were used as plant material. Agrobacterium tumefaciens EHA105 strain (Hood et al. 1993) was used, carrying a GUS-intron plasmid (Vancanneyt et al. 1990) as a transformation vector. Seeds previously disinfected, peeled and germinated for 24 hrs in MS1/2 media, were cut in half, resulting in 2 hemiembryos each one with a complete cotyledon. The cut was done inside a Petri dish with 20 ml of bacteria (OD660: 0.05) grown in AIM media supplemented with acetosyringone. A co-culture (CC media) was performed for 2-4 days in darkness at 22ºC. C explants were treated in the same way as A but co-culture treatment was performed in absence of Agrobacterium. For the regeneration step explants were transferred every 10/14 days through different regeneration media (Re1 to Re4) containing the antibiotic Tmt 0.3 mg/ml to inhibit Agrobacterium growth, as was previously determined in our laboratory. Finally, a passage to a rooting media for a 20 days period was made. In vitro conditions were 22-24ºC of temperature and 16 hrs light. Plantlets that showed a suitable radicular development were transferred to the greenhouse. Media formulation was based in Murashige and Skoog (1962). MS1/2: half saline concentration. All Re media contained: AgNO3 0.82 µg/l and Tmt 0.3 mg/ml. Re1: BAP 0.2 mg/ml and NAA 0.01 mg/ml. Re2: BAP 0.2 mg/ml. Re3: BAP 0.1 mg/ml. Re4: no hormones. RA (rooting): NAA 0.1 mg/ml and Tmt 0.3 mg/ml. Selection was performed from Re3; shoots C and A were divided in media with or without Km (50 mg/l). Note was taken from the phenotype of the shoots. Shoots that rooted in RA media with Km were transferred to the greenhouse. Χ2- Chi square statistical analysis was performed. GUS-intron gene expression was demonstrated by histochemical staining assays (Jefferson et al. 1987). Genomic DNA extraction was performed using the CTAB method (Saghai-Maroof et al. 1984). PCR was performed using Invitrogen Taq polimerase (1 U) and 0.1 µM of each primer (GUS391 UP: 5'-ACC GTT TGT GTG AAC AAC GA-3' and GUS2: 5'-GGC ATT GGT TTC GAA GCG GG-3'). Negative controls: mix without the addition of DNA and other with DNA from a non-transformed plant. Positive controls: GUS-intron plasmid DNA and DNA from Solanum tuberosum transformed with the uidA gene. A total of 724 plantlets were analyzed in 4 different culture situations: non-agroinfected controls (C), non-agroinfected controls treated with Km (C-Km), agroinfected (A) and agroinfected treated with Km (A-Km). In order to study the in vitro response to the antibiotic and determine if there were differential responses of the cultures in the 4 analyzed situations the following four parameters were studied:
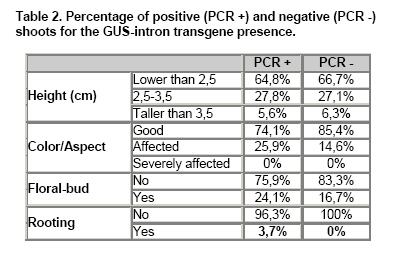
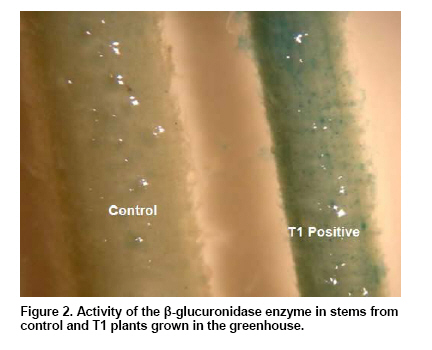
The results obtained are shown in Table 1. No significant differences were observed among the 4 analyzed treatments in none of the four studied parameters but interestingly no C-Km from 233 shoots was able to develop roots while in the group of A-Km, 2 shoots from 282 (0.7%) developed in vitro roots. These two shoots were transferred to the greenhouse and only one of them produced seeds (6 seeds) allowing the analysis of the T1 filial. The molecular analysis was done by PCR using the GUS-intron probes on the DNA of the following: a) 199 in vitro shoots from C, A y A-Km, b) leaves from the two plants transferred to the greenhouse that rooted in vitro in the presence of Km and c) leaves from the six plants grown in the greenhouse of the T1 filial. It must be taken into account that plantlets from a) and b) are chimeras. The PCR results, presence or absence of the transgene, of the A-Km shoots are summarized in Table 2. In order to find a relation between the molecular results and the observed in vitro shoot response, data from the A-Km shoots shown in Table 1 and in Table 2 were analyzed and compared. The statistical Χ2 analysis (p-value 0.1) revealed that there was no association between transgene presence and a given colour/aspect, height or the presence of floral-buds. In relation to in vitro rooting, as the number of rooted shoots was rather low no statistical difference was observed. Nevertheless, it must be highlighted that both rooted shoots in Km resulted positive by PCR analysis and that no control shoot could root in presence of the selective agent (Table 2). The T1 plants were studied by PCR and by histological GUS assay (Figure 2), resulting all of them positive. In previous sunflower transformation studies using Km selection for hemiembryos transformation at the same concentration that was used in this work, Knittel et al. (1994) found that the selection procedure induced discolouring of the sensible sectors while other resistant areas stayed dark green. The authors mentioned that these areas were transferred to a rooting media, obtaining 7% efficiency but no data on the escape level was presented. Malone-Schonenberg et al. (1994) reported a 20% escapes on selecting with 200 mg/l Km, and this level increased with 25 mg/l Km. Burrus et al. (1996) using two kinds of explants (complete apical embryonic meristems or half-cut) and 50 mg/l Km, observed 18% of escape while Lucas et al. (2000) reported a 72%. In order to avoid grafting difficulties, Hewezi et al. (2002), treated good colour and healthy shoots with a rooting solution in their base and the level of escapes was of 10%. On the other hand, Müller et al. (2001), performing a selection on regenerated shoots from a non-meristematic tissue with 1, 3, 10 and 15 mg/l Km, were not able to select transformed shoots and identify the transgenic events by Southern or Western analysis. In the present study the results obtained showed that the colour and aspect of the shoots are not valid physiological parameters for identification of transgenic events as 67.4% of the A-Km putative transformed plantlets according to these parameters are escapes (Table 1). Furthermore, 85.4% of the shoots from the non-transformed group (PCR -) presented a healthy aspect, so they would have been selected as putative transgenic (Table 2). In this way, it is shown here that there is no association between shoot colour and the presence of the transgene, therefore, this physiological condition must not be considered as a selection criterion in sunflower transformation. There is only one article with a different selection criterion, Grayburn and Vick (1995) reported that non-transformed shoots are unable to develop roots if they were cultured previously in a Km regenerating media. Up to now, our work is the first one where the selection criterion is the in vitro root development in presence of Km. Concluding RemarksThe selection by in vitro rooting in a Km culture medium is a promising successful method as no escapes were obtained and the transformation efficiency was of 0,7%. Another interesting conclusion is that shoot colour must not be considered as a selection criterion when Km is the selective agent in sunflower transformation.
Note: Electronic Journal of Biotechnology is not responsible if on-line references cited on manuscripts are not available any more after the date of publication. © 2006 by Pontificia Universidad Católica de Valparaíso -- Chile The following images related to this document are available:Photo images[ej06108f2.jpg] [ej06108t2.jpg] [ej06108f1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}