 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Electronic Journal of Biotechnology, Vol. 10, No. 1, Jan 15, 2007, pg. 1-8 RESEARCH ARTICLE Growth of Cunninghamella elegans UCP 542 and production of chitin and chitosan using yam bean medium Thayza Christina Montenegro Stamford1 , Tânia Lucia Montenegro Stamford2 , Newton Pereira Stamford3 , Benicio de Barros Neto4 , Galba Maria de Campos-Takaki*5 1Núcleo de Pesquisa
Financial support: Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq), Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES), and the Universidade Católica de Pernambuco (UNICAP). Code Number: ej07006 Abstract Microbiological processes were used for chitin and chitosan productions by Cunninghamella elegans (UCP 542) grown in a new economic culture medium. The assay was carried out to evaluate the growth of C. elegans using yam bean (Pachyrhizus erosus L. Urban) medium, in different times of growth (24, 48, 72 and 96 hrs), incubated at 28ºC in an orbital shaker at 150 rpm. The lyophilized biomass was determined by gravimetry. The polysaccharides were extracted by alkali-acid treatment, and characterized by infrared spectroscopy, titration and viscosity. C. elegans grown in the yam bean medium and produced higher yields of biomass (24.3 g/ mL) in 96 hrs. The high level was chitosan (66 mg/g), and chitin (440 mg/g) were produced at 48 and 72 hrs of growth, respectively. The polysaccharides showed degree of deacetilation and viscosimetric molecular weight as: 6.2% and 3.25 x104 g/mol for chitin, and 85% and 2.72 x 104 g/mol for chitosan, respectively. The results obtained suggest high biotechnological potential of yam bean as an economic source to produce chitin and chitosan by C. elegans. In addition, the new medium using yam bean for production of the chitin and chitosan may be used for many purposes to reduce the cost price of fermentation processes. Keywords: biopolymers, chitin, chitosan, Cunninghamella elegans, Zygomycetes.
Chitin, the insoluble linear β1,4- linked homopolymer of N-acetyl-D-glucosamine (GlcNAc), is the second most abundant natural polysaccharide (after cellulose). Chitosan is a cationic amino polysaccharide, essentially composed of β-1,4 D-glucosamine (GlcNAc) linked to N-acetyl-D-glucosamine residues (Andrade et al. 2000; Campos-Takaki, 2005), derived from de-N-acetylation of chitin (Tharanathan and Kittur, 2003; Amorim et al. 2005). These polysaccharides are found in a wide range of natural sources, such as crustaceans, insects annelids, molluscs, coelenterates and is a common constituent of fungal cell walls (Andrade et al. 2003; Synowiecki and Al-Khatteb, 2003; Franco et al. 2005). Chitin and chitosan hold a great economic value as due to their versatile biological activities and chemical applications, mainly in medical (Murugan and Ramakrishna, 2004; Yadav and Bhise, 2004) and pharmaceutical (Takeuchi et al. 2001; Kato et al. 2003) areas. The wide range of applications of these polymers has been extensively studied (DosSantos et al. 2003; Yusof et al. 2003; Campos-Takaki, 2005). Chitin and chitosan show peculiar properties, such as: biodegradability, biocompatibility, bioactivity, selective permeability, polieletrolic action, chelation, ion exchange properties, antitumor and antimicrobial activity (Dos Santos et al. 2003; Chung et al. 2004; Yadav and Bhise, 2004), and adsorption capacity (Shigemasa and Minami, 1996; Tharanathan and Kittur, 2003; Franco et al. 2004). Recent advances in fermentation technologies suggest that the cultivation of selected fungi can provide an alternative source of chitin and chitosan. The amount of these polysaccharides depends of the fungi species and culture conditions (Tan et al. 1996; Pochanavanich and Suntornsuk, 2002; Andrade et al. 2003; Synowiecki and Al-Khatteb, 2003). Filamentous fungi have been considered an attractive source of chitin and chitosan for industrial applications because their specific products can be manufactured under standardized conditions (Synowiecki and Al-Khatteb, 1997; Pochanavanich and Suntornsuk, 2002; Nemtsev et al. 2004). Usually, the Zygomycetes Class has higher amounts of chitin and chitosan in their cell walls when compared to other classes of fungi (Andrade et al. 2000; Campos-Takaki, 2005; Franco et al. 2005). The use of biomass from fungi has demonstrated great advantages, such as: independence of seasonal factor, wide scale production, simultaneous extraction of chitin and chitosan, extraction process is simple and cheap resulting in reduction in time and cost required for production, and also absence of proteins contamination, mainly the proteins that could cause allergy reactions in individuals with shellfish allergies (Andrade et al. 2000; Amorim et al. 2001; Nadarajah et al. 2001; Andrade et al. 2003; Franco et al. 2005). However, to optimize the production of chitin and chitosan from fungi, it's usually used complex or synthetics cultures media, which are expensive. It's becomes necessary to obtain economic culture media that promote the growth of fungi and stimulate the production of the polymers. Ecomonic microbial culture media normally use vegetables components. Yam bean (Pachyrhizus erosus L. Urban) is a leguminous plant native from the Amazon region and from Mexico semiarid region (Sorensen, 1996). The leguminous produces comestible tubercles and seeds with high level of protein and lipids. The tubercles were used as a good starch source for several industrial purposes (Sarangbin and Watanapokasin, 1999; Stamford et al. 1998; Stamford et al. 2001). The main characteristic of yam bean is the simple manipulation and low nutrition requirements when compared with other similar cultures, and tuberous roots yields is up to 60 t/ha (Stamford et al. 1998; Stamford et al. 2001; Stamford et al. 2003). The present paper aims to investigate the chitin and chitosan production using the Mucoralen fungi Cunninghamella elegans (UCP 542), grown by submerse fermentation in economic culture medium, yam bean (Pachyrhizus erosus L. Urban), as substrate, and was compared with four traditional culture media. Cunninghamella elegans UCP 542 (Culture Collection of Catholic University of Pernambuco,Recife , Brazil ) isolated from mangrove sediment situated in Rio Formoso, PE, Brazil . The strain was maintained at 4ºC on Potato Dextrose Agar (PDA) slants. C. elegans was grown, for chitin and chitosan production, in five different culture media: a) Sabouraud sucrose- (bacteriological peptone ( 10 g ) and sucrose ( 20 g ) per litre of distilled water, pH 5.7); b) Hesseltine and Anderson- (glucose ( 40 g ); asparagine ( 2 g ); chloridrate of thiamine (0.05 mg); potassium phosphate ( 0.50 g ) and magnesium sulphate ( 0.25 g ) per litre of distilled water, pH 5.2); c) Andrade et al. (2000)- (glucose ( 60 g ); asparagine ( 3 g ); chloridrate of thiamine (0.08 mg); potassium phosphate ( 0.50 g ) and magnesium sulphate ( 0.25 g ) per litre of distilled water, pH 5.1); d) Malte glucose- (Yeast Malte ( 17 g ) and glucose ( 20 g ) per litre of distilled water, pH 6.0); and e) Yam Bean Medium (Pachyrhizus erosus L. Urban)- basic chemical composition (total protein ( 8.72 g ), Starch ( 40.9 g ) and glucose ( 11.14 g ) per litre of distilled water, pH 7.0). Tuberous roots of yam bean were provided by the Department of Agronomy at Federal Rural University of Pernambuco (Northeast Brazil ). The tuberous roots surface were washed with water and soap, for withdrawal of impurities. The Yam Bean Medium was prepared from tuberous roots peeled and rounded sliced (± 1,5 cm ) and boiled in distilled water in the ratio of 1:2 (w/v) for 45 min (after initiate the boil). The broth was cooled, filtered in filter of paper and autoclavated at 121, 5ºC, 15 min. Growth profile. The sporangioles of C. elegans were harvested from cultures grown for seven days at 28ºC on Petri dishes containing PDA medium. A suspension was prepared and adjusted to 108 sporangioles/mL, using a hematocytometer for counting. For fungal submerse cultivation, 10 mL sporangioles suspension (108 sporangioles/mL) were inoculated in Erlenmeyer flask of 1000 mL containing 290 mL of culture media, and the flasks were incubated at 28ºC in an orbital shaker at 150 rpm, during 96 hrs. The mycelia were harvested, washed twice in distilled and deionised water by filtration, utilizing a silkscreen nylon membrane ( 120 F ), and were submitted to lyophilization process. After lyophilization the biomass was maintained in a vacuum dissecator until constant weight. During C. elegans submerse cultivation in yam bean medium, aliquots were collected every 24 hrs for biomass, pH, glucose and total nitrogen determination. Glucose
and nitrogen consumption and pH determination. The glucose consumption
was determined by the enzymatic colorimetric method (Labtest® Kit - Glucose oxidase). A standard curve was made by using a range
of glucose solutions (0.5 to 10.0 g/L). Colorimetric method Labtest® Kit for protein was utilized for the nitrogen consumption determination,
using a spectrophotometer Spectronic Genesys 2. Changes in pH were
measured using a potentiometer (Digital Pontentiometer Quimis Mod.
Chitin and chitosan extraction. The process of extraction involved deproteination with 2% w/v sodium hydroxide solution (30:1 v/w, 90ºC , 2 hrs), separation of alkali-insoluble fraction (AIF) by centrifugation (4000 rpm, 15 min), extraction of chitosan from AIF under reflux (10% v/v acetic acid 40:1 v/w, 60ºC , 6 hrs), separation of crude chitin by centrifugation (4000 xg, 15 min) and precipitation of chitosan from the extract at pH 9.0, adjusted with a 4 M NaOH solution. Crude chitin and chitosan were washed on a coarse sintered-glass funnel with distilled water, ethanol and acetone and air-dried at 20ºC (Franco et al. 2004). Chitin and chitosan characterization Infrared spectroscopy (Deacetylation degree - DD%). The degree of deacetylation for microbial chitin and chitosan were determined using the infrared spectroscopy according to Roberts (1992), using the absorbance ratio A1655/A3450 and calculated according to equation 1:
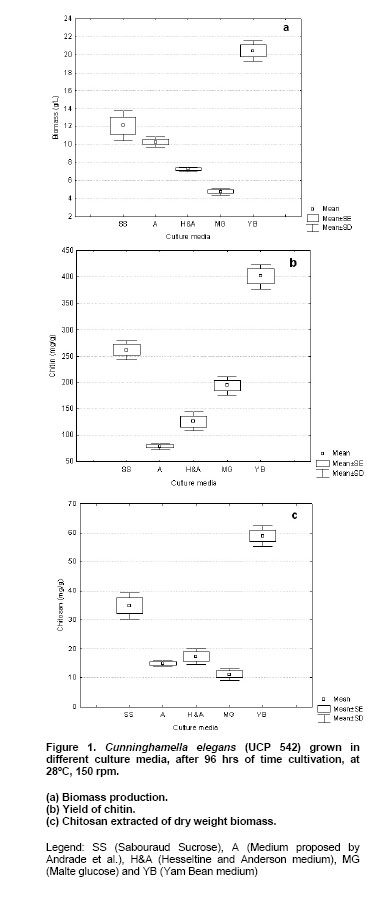
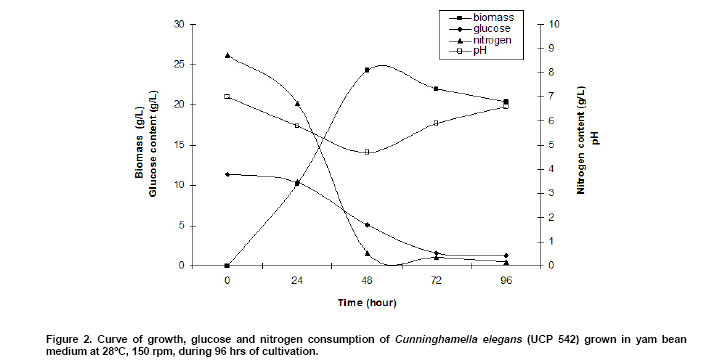
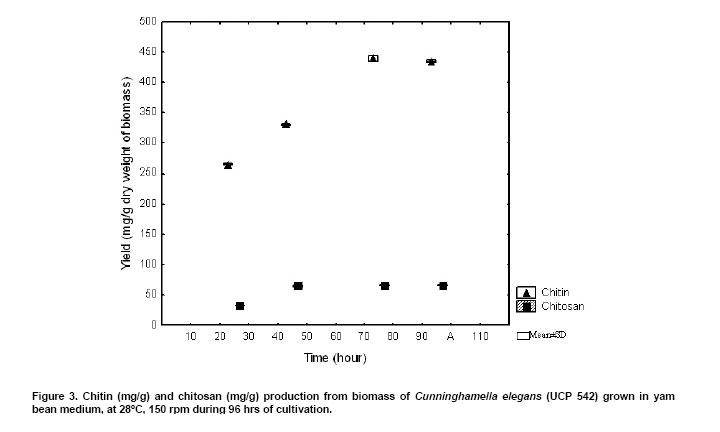
Two milligrams sample of fungal chitin and chitosan, which had been dried overnight at 60ºC under reduced pressure were thoroughly blended with 100 mg of KBr, to produce 0.5 mm thick disks. The disks were dried for 24 hrs at 110ºC under reduced pressure. Infrared spectrometer was recorded with a Bruker 66 Spectrometer, using a 100 mg KBr disks for reference. The intensity of maximum absorption bands were determined by the baseline method. Molecular weight. The molecular weights of chitin and chitosan were determined by viscosity, using the procedure described by Dos Santos et al. (2003). The viscosity of chitosan was determined using an AVS-350 viscometer (Schott-Geräte), type/capillary: Cannon-Fenskedinside= 1.01 mm , at 25ºC . After getting the intrinsic viscosity from tables K and a, were obtained for HAc/NaAc. K = 0.076, a = 0.76. The flow time was determined in seconds. Using Mark-Houwinks equation the average viscosimetric molecular weight expressed in g/mol. See equation 2. 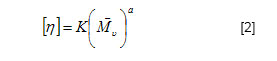 The data were analyzed for significance using the Student's t-test and chi-square test using STATISTICA program version 6.0 of Statsolt Inc., USA . All experiments were carried out in triplicate and the results are expressed as mean ± S.D. The profile of growth of C. elegans (UCP 542) and chitin /chitosan production using yam bean medium and four different culture media, traditional for Mucorales, are showed in Figure 1 . A higher production of biomass, statistically significant(p = 0.0028) can be verified in the yam bean medium, with average dry weight corresponding to 20.4 g/L (Figure 1a ). In addition, the best yield of chitin and chitosan per 1 g of biomass from C. elegans (UCP 542) are obtained using yam bean medium and Sabouraud sucrose medium for chitin 400.9 mg/g and 261,7 mg/g and chitosan 58,9 mg/g and 34,9 mg/g, respectively (Figure 1b and Figure 1c ). The results are superior to the reported by Andrade et al. (2000) and Franco et al. (2005) which proposed C. elegans as a promising chitin source, obtained as higher chitin yields 280 mg/g and 240 mg/g, respectively. Andrade et al. (2003) reported a maximum chitin yield of 239 mg/g from Mucor javanicus. However, C. elegans (UCP 542) grown in Hesseltine and Anderson medium shows a biomass yield of 10.3 g/L, with 96 hrs of cultivation. The result is in agreement with the growth curve of C. elegans (IFM 46109) established by Andrade et al. (2000) and Franco et al. (2005) using the same culture medium, which referred biomass yield of 11.0 and 11.6 g/L, respectively. C. elegans (UCP 542) grown in malt medium with glucose 2% during 96 hrs, shows average biomass production of 4.8 g/L. This result is similar to the reported by Synowiecki and Al-Khatteb (1997) which obtained a yield biomass of Mucor rouxii grown in yeast extract and glucose 2% medium, for 48 hrs, to the 4 g , per litre of medium. Chitin and chitosan production by C. elegans grown in yam bean medium are studied on the cultivation profile. C. elegans (UCP 542) growth curve for biomass, pH, nitrogen content and glucose consumption is present in Figure 2. Biomass production increase rapidly up to 48 hrs, reached about the maximum of 24.3 g/L of dry weight. The result is superior to the value 10.41 g/L and 11.6 g/L reported by Andrade et al. (2000) and Franco et al. (2004), respectively, for C. elegans (URM 46109) grown during 96 hrs in Mucorales medium. The results in Figure 2, demonstrate the residual glucose and nitrogen were 1.27 g/L and 0.18 g/L respectively. Similar results were reported by Andrade et al. (2000) and Franco et al. (2004). Amorim et al. (2001) suggested that the remaining glucose and nitrogen as due to nitrogenous compounds like secondary metabolites present at the end of growth of the fungi metabolism and excess of carbon source in the medium. The pH of yam bean medium oscillates between 7 and 5, during the culture time (Figure 2). The values of pH decrease during the exponential phase, probably, by pyruvic acid formation, as due to the high glucose and starch concentration in yam bean medium. Franco et al. (2004) described constant values of pH during the "lag" phase and pH decrease during the exponential phase. That information of higher amount of the yam bean glucose and starch was previously mentioned by Sarangbin and Watanapokasin (1999), during the citric acid production. Amorim et al. (2001) reported that during growth of C. elegans the pH of the media drops in the first 24 hrs and remained low (between 3 and 4) during the first 96 hrs of cultivation, probably because of the interaction between the medium substrate and the release of ions from the cell. Figure 3 shows chitin and chitosan yield extracted, to each 24 hrs, from C. elegans (UCP 542) grown in yam beam medium during 96 hrs of cultivation. The best yields of the polysaccharides (mg per gram of dry mycelia biomass) are obtained with 48 hrs of culture for chitosan (66 mg/g or 6.6%) and with 72 hrs for chitin (440 mg/g or 44%). Similar results were reported to Tan et al. (1996), which studied different Zygomycetes strains and observed that Cunninghamella echinulata was the best chitosan producing strain, with a yield of approximately 7.0% of chitosan per mycelia dry weight. Chitosan production stabilize after 48 hrs of culture, although, chitin production increase up to 72 hrs of culture, and decrease at 96 hrs. The higher chitosan yields in 48 hrs of growth suggest that during initial growth chitin is less crystalline and thus more susceptible to chitin deacetylase, and the chitosan formed by this enzime prevails in acid pH (Figure 3). According to Amorim et al. (2005) optimum pH for chitin deacetylase activity from Zygomycetes is pH 4.5. During C. elegans growth the pH of the yam bean medium drops in the first 48 hrs (Figure 2), stowing high metabolic interchange between the medium substrate uptake and the release of ions from the cells. Amorim et al. (2001) reported higher yields of chitosan were found within 24 hrs of cultivation of M. racemosus and C. elegans at pH 3.5, which seems to be also a stimulating agent for production of this biopolymer. The data in the present work are in agreement with Pochanavanich and Suntornsuk (2002) and Nadarajah et al. (2001) that chitosan production by the microorganisms is strongly dependent of the culture conditions, including cultivation time. Chung et al. (2004) demonstrated that chitin and chitosan content in the cellular wall of fungi change according to the species and these polymers usually show higher values in Zygomycetes. The characterization of chitin and chitosan obtained from C. elegans in yam bean medium by infrared spectra are similar to those reported in the literature (Andrade et al. 2000; Amorim et al. 2001; Franco et al. 2005). The most significant parts of chitin and chitosan spectra are those showing the amide bands at approximately 1665, 1555 e 1313 cm-1, which could be assigned to the C = O stretching, the N-H deformation in the CONH plane and the CN bond stretching plus CH2 wagging. In a similar way, chitin from C. elegans shows bands in the amide II region, which were 1153, 1378 and 1558 cm-1. The results are in agreement with Shigemasa and Minami (1996). Andrade et al. (2003) and Franco et al. (2005) which reported that chitin structure containing two types of amide group and both form C = O N-H intermolecular bonds, but one is also an acceptor for the CH2OH group. According to Dos Santos et al. (2003) the deacetylation and the regeneration process, cause disturbance in the initial crystalline reticulum of chitin, inducing a reordering of the hydrogen linking of chitosan. This can be observed in the central band at approximately 3483 cm-1 e 3305 cm-1, in the region of the axial deformation of OH, which appears overlapping the band of axial deformation of NH indicating the intermolecular hydrogen linking formation, and at the displacement of the higher frequency band indicating an increase in the structural order. The data are in accordance with the reported in literature when comparing both chitin and chitosan infrared spectra obtained by microbiological methods (Andrade et al. 2000; Amorim et al. 2001; Pochanavanich and Suntornsuk, 2002; Dos Santos et al. 2003; Franco et al. 2005). Deacetylation degree (%DD) is an important parameter associated with the physical-chemical properties of chitosan, because it's linked directly to the chitosan cationic properties (Pochanavanich and Suntornsuk, 2002). In the present study chitin and chitosan from C. elegans grown in yam bean medium present 6,2% DD and 85% DD, respectively. Amorim et al. (2001), Pochanavanich and Suntornsuk (2002) and Franco et al. (2004), reported deacetylation degree of chitosan from fungi between 80 to 90% DD. The average viscosimetric molecular weight (MV) of chitin and chitosan from C. elegans obtain in the present research are 3.25 x 104 g/mol for chitin, and 2.72 x 104 g/mol for chitosan. The results are in agreement with the results found in the literature, molar weights ranged between 1,0 x 104 to 9,0 x 105 g/mol (Nadarajah et al. 2001; Pochanavanich and Suntornsuk, 2002; Dos Santos et al. 2003). The results present here suggest a high biotechnological potential of yam bean as an economic medium to chitin and chitosan production by C. elegans, and may be used to reduce the cost price of these polysaccharides production.
Note: Electronic Journal of Biotechnology is not responsible
if on-line references cited on manuscripts are not available any more after
the date of publication.
© 2007 by Pontificia Universidad Católica de Valparaíso -- Chile The following images related to this document are available:Photo images[ej07006f3.jpg] [ej07006f1.jpg] [ej07006f2.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}