 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Electronic Journal of Biotechnology, Vol. 10, No. 1, Jan 15, 2007, pg. 1-9 RESEARCH ARTICLE Influence of 2,4-D and BAP on callus growth and the subsequent regeneration of somatic embryos in long-term cultures of Pelargonium x domesticum cv. Madame Layal Klaus-Thomas Haensch1
1 Institute
of
Vegetable
and Ornamental Crops Großbeeren/Erfurt e.V.,
Department Plant Propagation,
Kühnhäuser Str. 101, D-99189 Erfurt-Kühnhausen,
Germany
Tel: 49 0 36 201 785 224
Fax: 49 0 36 201 785 250
E-mail: haensch@erfurt.igzev.de Financial support: This investigation was undertaken with the support of the Ministries of Agriculture of the Federal Republic of Germany and the States of Brandenburg and Thüringen. Code Number: ej07007 Abstract This long-term study demonstrates for the first time that it is possible to propagate embryogenic cultures in pelargoniums and to subsequently initiate the differentiation of embryos using the cultivar Madame Layal (Pelargonium x domesticum). Propagation of callus was only possible with combinations of 2,4-dichlorophenoxyacetic acid (2,4-D) and 6-benzylaminopurine (BAP), which gave rise to embryos from the primary culture stage on. However, the propagation of cells, as well as the differentiation of embryos, was inhibited by a continuous application of these growth regulators. For this reason, a long period on medium lacking growth regulators was necessary before the differentiation of embryos occurred again. The consequences for improving the propagation of embryogenic cultures in pelargoniums are discussed. Keywords: auxin, cytokinin, pelargonium, propagation, somatic embryogenesis.
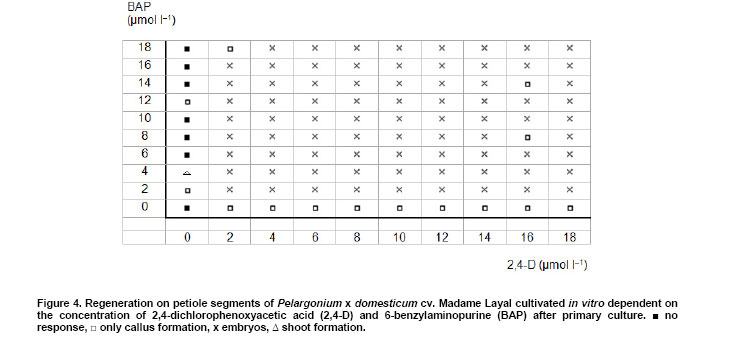
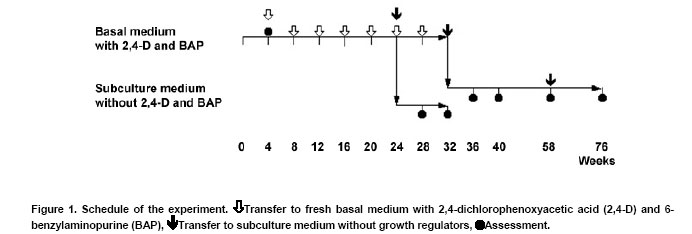
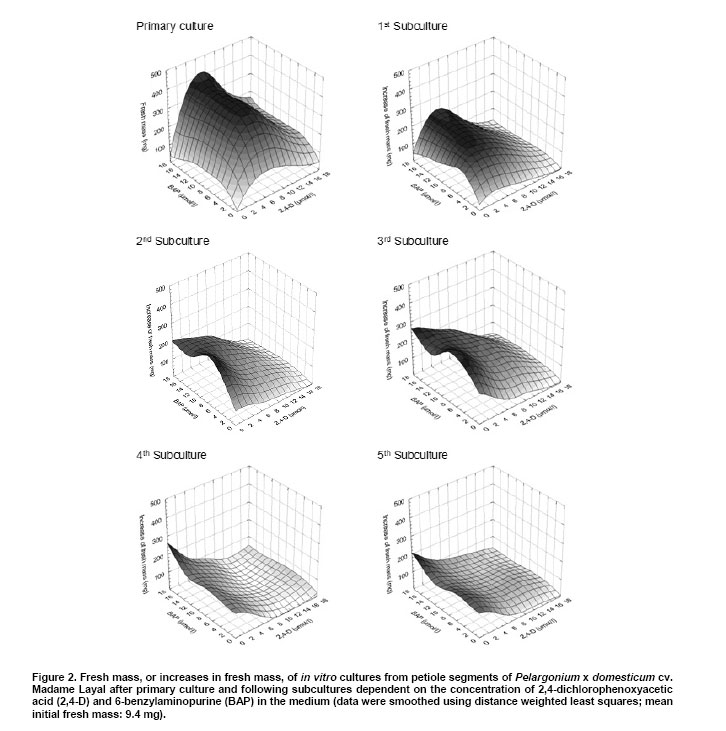
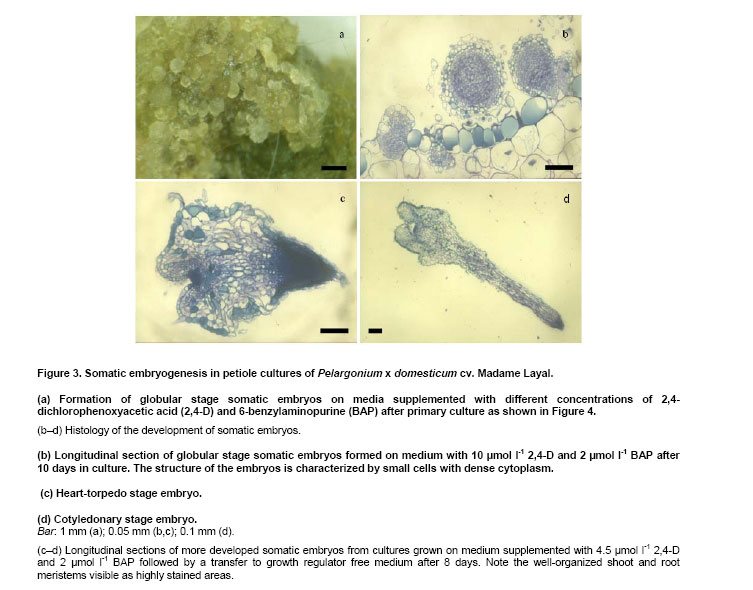
Pelargoniums are very popular ornamental plants, and their mass propagation is performed using cuttings or seeds (Mithila et al. 2001), which is very expensive. Somatic embryogenesis has the potential for establishing more efficient propagation systems. Most published studies on somatic embryogenesis in ornamental pelargoniums are based on explants, in particular hypocotyls, of very young seedlings of Pelargonium x hortorum (Marsolais et al. 1991; Hutchinson and Saxena, 1996a; Hutchinson and Saxena, 1996b; Hutchinson et al. 1996a; Hutchinson et al. 1996b; Murthy et al. 1996; Wilson et al. 1996; Croke and Cassells, 1997; Hutchinson et al. 1997a; Hutchinson et al. 1997b; Murthy et al. 1999; Senaratna et al. 1999; Hutchinson et al. 2000; Madakadze and Senaratna, 2000; Madakadze et al. 2000; Murch and Saxena, 2001; Senaratna et al. 2002). After a critical evaluation of these published results, and a consideration of morpho-histological studies of somatic embryo-like structures from their cultures, it was concluded that there has not yet been any proof of somatic embryogenesis from hypocotyls of this species (Haensch, 2004a). A repeated examination of this question by Madden et al. (2005) did not change this situation because once again no root pole formation could be demonstrated. However, "the most distinctive characteristic of an embryo is its anatomically discrete (closed) radicular end" (Haccius, 1978). Other studies are based on petioles. In Pelargonium x hortorum, "vegetative embryos" were detected in callus formed from petioles using a medium containing zeatin and α-naphthaleneacetic acid, and then a medium containing zeatin as the only growth regulator (Cassells, 1979). In the same species, the formation of "putative somatic embryos" in petiole cultures has been reported after applying thidiazuron (Croke and Cassells, 1997). In Pelargonium peltatum hybrids, there is a report about the regeneration of somatic embryos from petiole-derived suspension cultures using a medium with α-naphthaleneacetic acid and thidiazuron followed by a transfer to a medium with only thidiazuron and a subsequent transfer to a medium without plant growth regulators (Robichon et al. 1997). There is another report about the regeneration of somatic embryo-like structures from petioles using 6-benzylaminopurine (BAP) or thidiazuron in different combinations with indole-3-acetic acid or indole-3-butyric acid (Wojtania et al. 2004). In all these reports, the classification of regenerating structures as somatic embryos is unconvincing. This is because, according to the criteria formulated by Haccius (1978), the presented structures have not been shown to be clearly separated from parental explants nor have the more developed structures shown to be real bipolar structures, i.e., structures with a shoot and a root pole. The possibility of inducing somatic embryogenesis using thidiazuron in petiole cultures of Pelargonium x hortorum and Pelargonium x domesticum was histologically examined. It revealed that no somatic embryos were formed, and regenerants had to be classified as shoots and shoot- or leaf-like structures (Haensch, 2004b). These findings are supported by Winkelmann et al. (2005) who also found only shoots after applying thidiazuron in petiole cultures of pelargoniums. In other regeneration experiments with petioles of Pelargonium x hortorum and Pelargonium x domesticum, the most effective treatment for the induction of somatic embryogenesis was a medium supplemented with a combination of 2,4-dichlorophenoxyacetic acid (2,4-D) and BAP (Marsolais et al. 1991). Histological examinations of the regeneration response after the application of this method to the cultivar Madame Layal (Pelargonium x domesticum) confirmed that somatic embryos were formed, i.e., bipolar structures (Wilson et al. 1994). Although this protocol in combination with this cultivar was very suitable for the production of somatic embryos (Marsolais et al. 1991; Haensch, 1999), the regeneration had already occurred within four weeks after culture initiation, even on the medium with 2,4-D and BAP. For this reason, petioles are always needed for this protocol as the responding material for each regeneration process. In other species, for example Daucus carota, it is possible to propagate undifferentiated embryogenic callus on a 2,4-D-containing medium on which the development of embryos is suppressed by auxin. Afterwards, the differentiation of embryos can be initiated by transferring the callus to a medium lacking auxin (Komamine et al. 1990). Until now, there has been no procedure available in pelargoniums that allows the continuous propagation of undifferentiated embryogenic material followed by the regeneration of somatic embryos. Therefore, the objective of this study was to examine to what extent such a procedure would be possible in this species. For this purpose, we investigated the connection between different amounts of auxin and cytokinin in the medium and the resultant growth and regeneration responses. The experiment was performed using petioles of the cultivar Madame Layal (Pelargonium x domesticum). Mother plants were grown in a growth chamber at 20ºC under a 16/8 hrs (light/dark) photoperiod with light supplied by white fluorescent lighting at an intensity of 46 µmol m-2 s-1. Petioles from young partially expanded leaves were surface-sterilized according to the method of Marsolais et al. (1991) by agitating them for 20 min in a 1.4% solution of sodium hypochlorite containing one drop of Tween 20 per 400 ml followed by five separate rinses with sterile distilled water. The petioles were cut into 5 mm long segments and plated side-down, regardless of the orientation in vivo, on the surface of the culture medium. Each explant was cultivated in a 50 ml test tube containing 10 ml medium. The test tubes were sealed with plastic film and incubated in a growth chamber at 24ºC under a 16/8 hrs (light/dark) photoperiod with light supplied by white fluorescent lighting at an intensity of 46 µmol m-2 s-1. MS medium (Murashige and Skoog, 1962) with 50 mg l-1 ascorbic acid, 50 mg l-1 citric acid (Marsolais et al. 1991), 30 g l-1 sucrose, and 6.5 g l-1 agar at pH 5.6 was used as the basal medium. Iron was provided as the ethylenediaminetetraacetic acid iron salt, and its content was lowered to 86.9 µmol l-1. This medium was supplemented with 2,4-D and BAP at concentrations between 0 and 18 µmol l-1 according to the experimental scheme (Figure 4). It was then autoclaved for 20 min at 121ºC . Each treatment included five explants, and was not repeated. Instead of replications, the experimental scheme was extended to hundred different combinations of growth regulators to allow a comprehensive assessment of responses. The primary culture was followed by five subcultures of four weeks each on the same medium (Figure 1). After this time, half of the material in each treatment was transferred to the same medium without growth regulators for eight weeks to check the ability of the grown callus to regenerate embryos. During transfer, all embryos that had already formed were removed. The second half of the material was maintained on the former medium for a further two subcultures and then transferred to growth regulator-free medium. After 26 weeks on this medium, the material was transferred to the same medium for a further 18 weeks. For growth characterization, the fresh mass of the cultures was measured after the primary culture and after each of the first five subcultures on the growth regulator-containing medium. In addition, the initial value of fresh mass was determined using petiole segments from a sample of 30 explants. The kind of regeneration was determined after primary culture, four and eight weeks after transfer to the growth regulator-free medium following the fifth subculture on growth regulator-containing medium, or four, eight, and 26 weeks after transfer to the growth regulator-free medium following the seventh subculture on growth regulator-containing medium (Figure 1). A further assessment of regeneration was performed 18 weeks after the last transfer of the material to medium without 2,4-D and BAP. The regeneration of somatic embryos on media with 2,4-D and BAP was examined histologically. For this purpose additionally cultivated petioles with globular somatic embryos were used after 10 days of the primary culture from the treatment with 10 µmol l-1 2,4-D and 2 µmol l-1 BAP. More developed somatic embryos were obtained by cultivating petiole explants by the same methods as mentioned above with the exception that GCM medium solidified with 6 g l-1 agarose and supplemented with 4.5 µmol l-1 2,4-D and 2 µmol l-1 BAP was used followed by a transfer to the same medium without growth regulators after eight days (Marsolais et al. 1991) to increase the number of available structures. Altogether 24 explants with developing globular somatic embryos and seven more developed somatic embryos were examined. The histological examinations were performed as described by Haensch (2004a). The toluidine blue stains the cytoplasm and unlignified cell walls red and the DNA-containing structures and lignified cell walls blue. The cultures produced different amounts of fresh mass dependent on the growth regulator combination and subculture (Figure 2). Based on the missing homogeneity of variances between the treatments, no analysis of variance could be performed (primary culture:Bartlett test χ2 = 247.4; p < 0.01). For this reason, the dependence of the fresh mass, or rather the increase in fresh mass, on the auxin and cytokinin concentration was calculated using the method of distance weighted least squares. Figure 2 shows that the region with the highest fresh mass after primary culture is with 6-8 µmol l-1 2,4-D in combination with 6-18 µmol l-1 BAP. In the following five subcultures, a shift in the growth maximum towards lower 2,4-D concentrations could be observed (Figure 2). From the second subculture on, although this growth maximum seems to be mathematically below 2 µmol l-1 2,4-D, real values exist only from 2 µmol l-1 upwards because many cultures without auxin had died at each step. For BAP, such a shift in the growth maximum is less clear. From the third subculture on, the highest increase in fresh mass is situated in the region between 8 and 14 µmol l-1. Considering the complete experiment, the maximal growth decreases from the primary culture to the following subcultures. Most growth regulator combinations have already given rise to the formation of globular embryos during primary culture (Figure 3a and Figure 4). The histological examinations revealed that the globular stage somatic embryos are new individuals, which consist of small cells with a dense cytoplasm (Figure 3b) and develop to bipolar structures showing clearly well developed shoot and root meristems (Figure 3c and Figure 3d). Combinations without 2,4-D showed no somatic embryogenesis. The explants often died or remained unchanged. Some of the explants formed a small callus. The combination of 0 µmol l-1 2,4-D with 4 µmol l-1 BAP gave rise to shoot regeneration. Combinations without BAP also showed no somatic embryogenesis, but small callus formed. All treatments with both BAP and 2,4-D gave rise to somatic embryogenesis. Exceptions were for the treatments 2 µmol l-1 2,4-D with 18 µmol l-1 BAP and 16 µmol l-1 2,4-D with 8 or 14 µmol l-1 BAP (Figure 4). A check on the ability to regenerate somatic embryos on growth regulator-free medium after five and seven subcultures could still be performed for nearly all combinations. In treatments without auxin, this was restricted to combinations with 2, 4, and 12 µmol l-1 BAP. A check after the fifth subculture showed that there were no somatic embryos four weeks after transfer to growth regulator-free medium. Only in some combinations were a few embryos found after remaining on this growth regulator-free medium for an additional four weeks without transfer to fresh medium, i.e., only between one and three embryos on one explant each of the 2,4-D/BAP treatments 2/12, 2/14, 4/2, 8/16, 10/10, 10/18, and 14/4 µmol l-1. A check after the seventh subculture on growth regulator-containing medium demonstrated that, after a further four and eight weeks on growth regulator-free medium, there were no somatic embryos. No somatic embryos were observed, even 26 weeks after transfer to growth regulator-free medium. Therefore, most of this material was discarded after this stage. Only one or two calluses of the treatments 2/4, 2/8, 2/12, 2/14, 2/18, 4/10, 4/18, 8/4, 8/12, and 10/10 µmol l-1 2,4-D/BAP were transferred again to fresh growth regulator-free medium. With all of these maintained combinations on calluses that were clearly grown during this time on this medium, somatic embryos with the same appearance as in the primary culture (Figure 3a) were found after 18 weeks. Their number varied between 1 and 22. Until now, no procedure could be established in pelargoniums in which, after a continuous propagation of undifferentiated embryogenic material, a specific stimulus would trigger the regeneration of somatic embryos. Although there is a report about the regeneration of somatic embryos from suspension cultures in P. peltatum hybrids (Robichon et al. 1997), the classification of regenerating structures as somatic embryos is unconvincing because it was not shown that the one presented structure was clearly separated from the parental tissue and that the other more developed structures were real bipolar structures. The only histological report clearly confirming the formation of bipolar structures in pelargoniums is the one involving somatic embryogenesis in the cultivar Madame Layal (Pelargonium x domesticum) (Wilson et al. 1994). The results of this report are confirmed by the present study. The regenerated somatic embryos developed as new individuals that consisted in the globular stage of the typical small cells with a dense cytoplasm and developed to clear bipolar structures (Figure 3b,c and d). The regeneration of somatic embryos in this cultivar is based on the protocol of Marsolais et al. (1991), which used petioles as explants and applied a medium with 2,4-D and BAP. In this protocol, the regeneration of somatic embryos had already appeared within four weeks after culture initiation, and even on the medium with 2,4-D and BAP. Therefore, possibilities for somatic embryogenesis in pelargoniums are restricted at present to the production of embryos on primary explants. In other species, for example D. carota, undifferentiated embryogenic callus can be established and propagated on a 2,4-D-containing medium, whereas a subsequent transfer to a medium lacking auxin initiates the differentiation of embryos (Komamine et al. 1990). Therefore, the objective of this study was to examine to what extent such a method would be realizable in pelargoniums. For this purpose, we investigated the connection between different concentrations of 2,4-D and BAP in the medium and the resultant growth and regeneration responses of petiole cultures of the cultivar Madame Layal (Pelargonium x domesticum) in a long-term experiment. The results represent a comprehensive survey of these responses, which were obtained with a small sample size and without a replication, but with a high number of growth regulator combinations. This special statistical approach is suitable to get such a survey (Baetz et al. 1982) but prevents to make differences between single treatments statistically evident. This has to be considered when such special differences will be assessed. Concerning the growth of the cultures, it became apparent that growth at a combination of 10 µmol l-1 2,4-D with 2 µmol l-1 BAP, which Marsolais et al. (1991) found to be the optimum concentration for the regeneration of somatic embryos in cv. Madame Layal, was much lower than the maximum values at all culture stages (Figure 2). Furthermore, the maximum of fresh mass increase shifted with subsequent subcultures from 6-8 µmol l-1 2,4-D in combination with 6-18 µmol l-1 BAP to very low 2,4-D concentrations in combination with almost unchanged BAP concentrations. An explanation for the considerably lower growth at higher auxin concentrations with increasing culture time could be growth inhibition caused by the enrichment of 2,4-D in the tissues, as was shown in Arabidopsis (Meijer et al. 1999). The regeneration observed during primary culture shows that, with this cultivar, the formation of globular embryos already occurs in a broad spectrum of 2,4-D/BAP combinations four weeks after culture initiation on these media. On the one hand, these embryos are produced even in situations in which the concentrations are suboptimal or already clearly inhibiting growth (Figure 2, primary culture, and Figure 4). On the other hand, in growth regulator combinations without the regeneration of embryos, the callus growth is so small that it is unsuitable for propagation. For this reason, propagation is only possible with combinations that trigger the regeneration of embryos already in the primary culture. However, our results show that, after five and seven subcultures on these media, embryogenic responses are rarely or no longer detectable if the cultures are transferred to growth regulator-free medium. Several explanations for this effect are possible. One possibility is that the medium without growth regulators might be unsuitable for the development of embryos. However, Marsolais et al. (1991) showed that the used cultivar regenerates embryos after a 2,4-D application of only 2-8 days followed by a transfer to growth regulator-free medium. Therefore, this explanation can be excluded. Another possibility is the embryos originate from already embryogenic determined cells. Such embryogenesis can lead directly to globular embryos through which the stock of embryogenic cells would be exhausted after a certain time. In the histological analysis of Wilson et al. (1994), no pre-embryogenic determined cells were reported. The analysis shows that normal sub epidermal parenchyma cells are the origin of embryogenesis. These cells are stimulated to embryogenesis by the application of 2,4-D in combination with BAP. For this reason, the greatly diminished embryogenic response in a growth regulator-free subculture following a longer application of 2,4-D and BAP cannot be explained by the exhaustion of a stock of embryogenic determined cells. It has to be interpreted as an inhibition effect of the long-lasting influence of the growth regulators used. This explanation is also supported by the fact that, in the case of very long subculture on growth regulator-free medium, some embryos are again formed on newly formed callus. The same processes as were discussed with regard to the inhibition of growth supposedly cause this inhibition. These results demonstrate for the first time that it is possible to propagate embryogenic cultures in pelargoniums, and to subsequently initiate the differentiation of embryos using the cultivar Madame Layal (Pelargonium x domesticum). However, the propagation of cells, as well as the differentiation of embryos, was inhibited for a long time by a continuous propagation on media containing 2,4-D and BAP. This knowledge could contribute to improving the propagation of embryogenic cultures and the subsequent regeneration of embryos in Pelargonium x domesticum. It could even be applied to the economically important Pelargonium x hortorum and P. peltatum hybrids, because using the protocol in these species gave rise to somatic embryos, but with only very low regeneration rates (Marsolais et al. 1991; Haensch, 1999). One approach for such improvements might be, for example, the elimination of growth regulators after a certain time by binding them to active charcoal or washing them away by frequently changing the medium. This has proven successful in overcoming the inhibition of somatic embryogenesis caused by accumulated 2,4-D in Arabidopsis cell suspensions (Meijer et al. 1999). Other approaches might be the repeated switching between development and disorganization of somatic embryos, as was found suitable for maintaining embryogenic competence in carrots (Nomura, 2003), or the substitution of growth regulators during the induction of somatic embryogenesis by alternative means such as salt stress, as was also possible in ginseng (Choi et al. 1998). The author
would like to thank Mrs Barbara
Note: Electronic Journal of Biotechnology is not responsible if on-line references
cited on manuscripts are not available any more after the date of publication.
© 2007 by Pontificia Universidad Católica de Valparaíso -- Chile The following images related to this document are available:Photo images[ej07007f3.jpg] [ej07007f1.jpg] [ej07007f4.jpg] [ej07007f2.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}