 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Electronic Journal of Biotechnology, Vol. 10, No. 1, Jan 15, 2007, pg. 1-14 RESEARCH ARTICLE Isolation and characterization of hydrocarbon producing green alga Botryococcus braunii from Indian freshwater bodies Chandrappa Dayananda1 , Ravi Sarada*2 , Vinod Kumar3 , Gokare Aswathanarayana Ravishankar4 1Plant
Cell Biotechnology Department,
Central Food Technological Research Institute,Mysore
570 020,
India
Tel: 91 821 2516501
Fax: 91 821 2517 233
E-mail: nanda.daya@gmail.com
Financial support: Department of Biotechnology, Government of India,New Delhi , India . Code Number: ej07008 Abstract Samples of green colonial unicellular microalga Botryococcus braunii were collected from Bear Shola Falls at Kodaikanal (latitude 10.31 N and longitude 77.32 E), Tamil Nadu, India . Specimens were isolated, cultured and examined for its hydrocarbon content, morphological features and DNA structural resemblance with the known strain to confirm its identity. Inter simple sequence repeats (ISSR) finger printing revealed strong genetic similarity among the authentic strain (B. braunii N-836) and the Indian isolated strain (B. braunii CFTRI- Bb1) from Kodaikanal. The type of hydrocarbons produced by the Kodaikanal isolates were analyzed and identified as saturated hydrocarbons in the range of C21 to C33 by GCMS. Tetracosane and octacosane were found as the major components among the saturated hydrocarbons produced by this alga, constituting 17.6% and 14.8% respectively. Hydrocarbon content of the organism was in the range of 13-18% of its dry biomass. The fat content of the organism was found to be 22% (w/w). Palmitic and oleic acids were found to be major fatty acids produced by the alga. Lutein and β-carotene were found to be the major carotenoids and constituted about 64.1% and 25.1% respectively of the total carotenoids. Based on ISSR finger printing and hydrocarbon analyses the new algal strain from Kodaikanal was identified as Botryococcus braunii. Keywords: biomass, carbon dioxide, carotenoids, fatty acids, inter simple sequence repeats, saturated hydrocarbons.
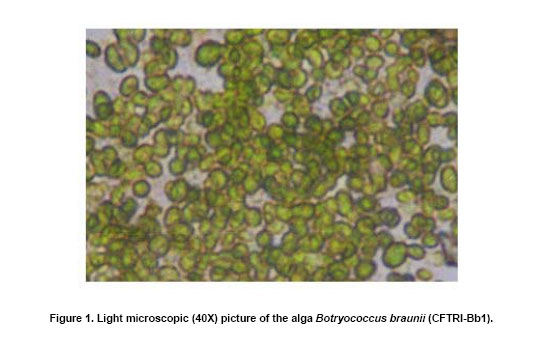
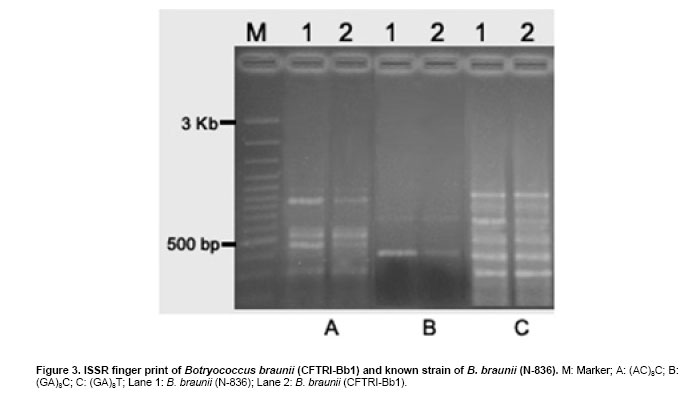
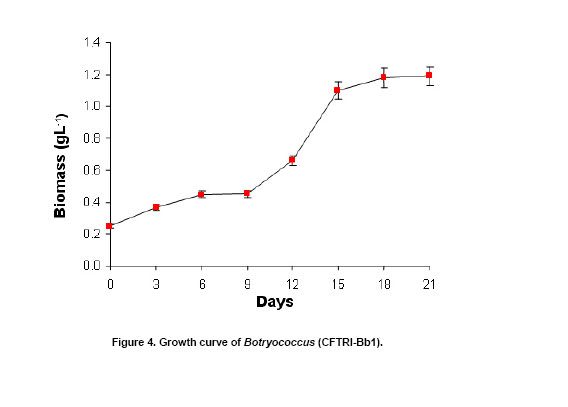
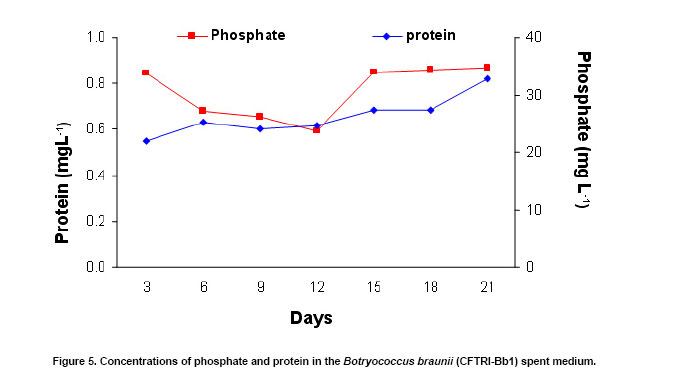
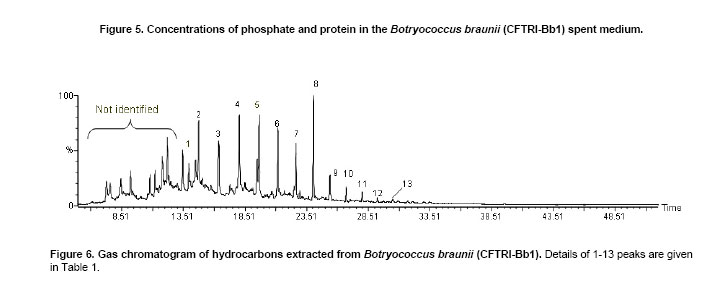
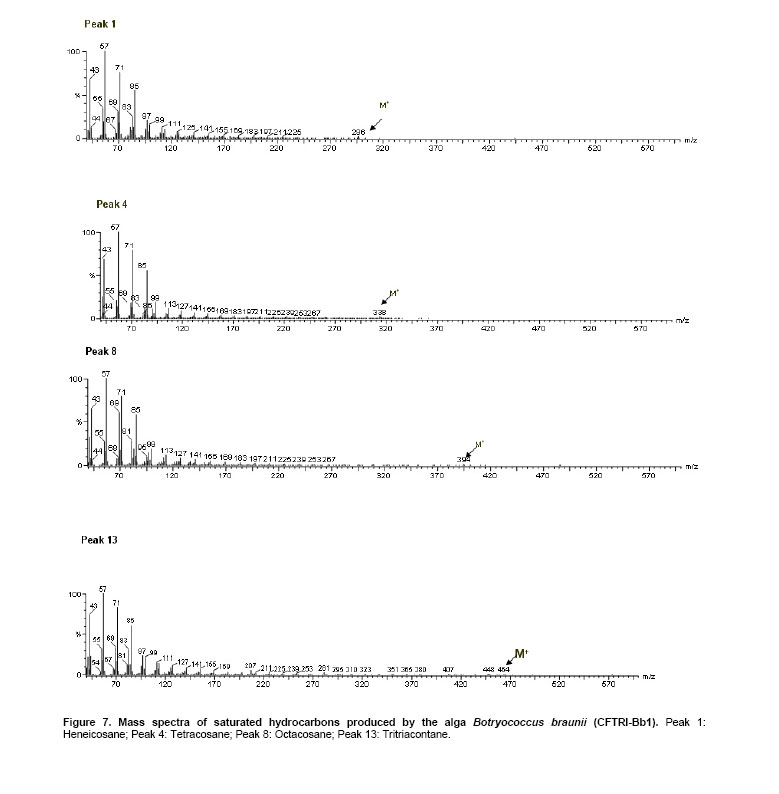
The green colonial hydrocarbon rich unicellular microalgae Botryococcus braunii (Banerjee et al. 2002; Metzger and Largeau, 2005) is widespread in freshwater, brackish lakes, reservoirs and ponds. It is also widely distributed in reservoirs at temperate, tropical and arctic latitudes (Tyson, 1995). It is recognized as one of the potent renewable resource for production of liquid hydrocarbons. B. braunii is classified into A, B and L races based on the type of hydrocarbons produced (Metzger and Largeau, 2005). Race - A produces C21 to C33 odd numbered n-alkadienes, mono-, tri-, tetra-, and pentaenes and they are derived from fatty acids (Banerjee et al. 2002; Metzger et al. 2005). The L race yields a single C40 isoprenoid hydrocarbon, lycopa -14(E),18(E)-diene (Metzger et al. 1990). The B race produces two types of triterpenes called botryococcenes of C30 - C37 of general formula Cn H2n-10 as major hydrocarbons and small amounts of methyl branched squalene. Certain strains of the B race also biosynthesise cyclobotryococcenes (David et al. 1988; Achitouv et al. 2004). Also a feature common to all three races is the presence of a highly aliphatic, non-hydrolysable and insoluble biomacromolecule (algaenan) found in their outer cell walls (Audino et al. 2002). The highly resistant nature of the B. braunii algaenan to degradation allows it to be selectively preserved during fossilisation, leading to fossil B. braunii remains, a major contributor to a number of high oil potential sediments (Simpson et al. 2003). The alga B. braunii produces hydrocarbon in the range of 2-86% (on dry weight basis). This variation in the content of hydrocarbon is due to the differences in the strains, the race it belongs and also depends on cultural and physiological conditions (Dayananda et al. 2005). Hydrocarbon oils extracted from B. braunii, when hydrocracked, produce a distillate comprising of 67% gasoline fraction, 15% aviation turbine fuel, 15% diesel fuel fraction and 3% residual oil (Banerjee et al. 2002; Dayananda et al. 2006), these fuels were free from oxides of sulphur and nitrogen (SOX and NOX) after their combustion. Being a photosynthetic organism, it can reduce CO2 emissions by 1.5 x 105 tons/yr and 8.4 X 103 ha of microalgal cultivation area would be necessary (Sawayama et al. 1999). The uptake of some toxic metals like chromium, cadmium and arsenic is also been reported (Sawayama et al. 1995). There is a need for isolation and identification of newer species or strains of Botryococcus which are efficient in hydrocarbon synthesis and could be adopted for the mass cultivation. In the present study focus was on isolation of indigenous strain of Botryococcus braunii from India and its characterization in hydrocarbons and carotenoids production. The effect of pH and carbon dioxide on its growth and hydrocarbon production was also studied and is reported here. An authentic strain of Botryococcus braunii (N-836) was obtained from the National Institute for Environmental Studies,Tsukuba , Japan . The stock culture was maintained on modifiedChu 13 medium. The samples were collected from different water bodies of Kodaikanal (latitude 10.31 N and longitude 77.32 E), ( India ) and cultured in modifiedChu 13 medium (Largeau et al. 1980). The algae were subjected to purification by serial dilution followed by plating. The individual colonies were isolated and inoculated into liquid medium (modifiedChu 13 medium) and incubated at 25 ± 1ºC under 1.2 ± 0.2 klux light intensity with 16:8 hrs light photoperiod. The purity of the culture was ensured by repeated plating and by regular observation under microscope. A time course study was carried out on B. braunii growth. The experiment was carried out in Erlenmeyer flasks of 150 ml capacity, containing 40 ml modifiedChu 13 medium for a period of 3 weeks. The culture flasks were inoculated (20% v/v) and incubated at 25 ± 1ºC under 1.2 ± 0.2 klux light intensity with 16:8 hrs light and dark cycle. Cultures were harvested and dry biomass was estimated at 3 days of intervals. Cell free medium was also analyzed for residual phosphate, protein content. All the experiments were carried out in triplicates. The effect of pH on growth of the alga and hydrocarbon yields was studied usingChu 13 media in the pH range of 6.0, 6.5, 7.0, 7.5, 8.0 and 8.5.The experiment was carried out in Erlenmeyer flasks (150 ml) containing 40 ml of Chu 13 modified medium and the pH of the medium was adjusted before autoclaving. All the flasks were inoculated uniformly at 25% (v/v) inoculum of 2 weeks old B. braunii culture. The culture flasks were incubated for 3 weeks at 25 ± 1ºC temperature with 1.2 ± 0.2 klux and 16:8 hrs light dark cycle, and the culture was harvested and analysed for biomass and hydrocarbon yields. A two- tier Erlenmeyer flask (Tripathi et al. 2001) was used for photoautotrophic growth experiments. The medium in upper chamber was inoculated with 25% (v/v) of two week old B. braunii culture. The mouth of the upper and lower compartments was sealed tightly with cotton plug and parafilm. The mixture of carbonate (3M) and bicarbonate (3M) solutions were added (100 mL) to the lower compartments to get a CO2 partial pressure of 0.5, 1.0 and 2.0% (v/v) respectively as given by Tripathi et al. (2001). The culture flasks were incubated for 3 weeks under 16:8 hrs light dark cycle with 1.2 ± 0.2 klux light intensity at 25 ± 1ºC . The experiment was carried out in triplicates. The cultures were harvested and analysed for biomass and hydrocarbon yields. The cultures were harvested and the cells were washed with distilled water after centrifugation at 5000 rpm. Then the pellet was freeze dried. The dry weight of algal biomass was determined gravimetrically and growth was expressed in terms of dry weight. A known volume of culture was centrifuged (8000 rpm) for 10 min and the pellet was treated with known volume of methanol and kept in water bath for 30 min at 60ºC . Absorbance of the pooled extracts was measured at 652 and 665 nm and chlorophyll (a + b) was estimated using Lichtenthaler equations (Lichtenthaler, 1987). A known quantity of algal dry biomass was homogenized and extracted repeatedly with acetone. The pooled extracts absorbance was read at 470 nm and total carotenoid contents were quantified according to Lichtenthaler (Lichtenthaler, 1987). The cells were harvested by centrifugation at 8000 rpm and the supernatant was analyzed for phosphate content by Fiske-Subbarao's method (Fiske and Subbarow, 1925). Protein content in the cell free medium was analyzed byBradford protein assay (Zor and Selinger, 1996) Hydrocarbon was extracted in hexane after homogenizing the dry biomass in a mortar and pestle in the presence of glass powder and the supernatant recovered after centrifugation was evaporated to complete dryness under the stream of nitrogen. Hydrocarbon content was measured gravimetrically and expressed as dry weight percentage (Dayananda et al. 2005; Dayananda et al. 2006). Hydrocarbon extract was purified by column chromatography on silica gel. The hydrocarbon samples were analyzed on SPB-1 column ( 30 m x 0.32 mm ID x 0.25 µm film thickness) using GCMS equipped with FID and were identified by comparing their fragmentation pattern with standards (Sigma) and also with NIST library (Dayananda et al. 2005). Lipids were extracted with chloroform - methanol (2:1) and quantified gravimetrically. The fatty acid methyl esters (FAME) were prepared as per the procedure of Christie (1982). FAME were analyzed by GC-MS (PerkinElmer, Turbomass Gold, Mass spectrometer) equipped with FID using SPB-1 (poly(dimethysiloxane)) capillary column ( 30 m x 0.32 mm ID x 0.25 µm film thickness) with a temperature programming 130ºC to 280ºC at a rate of 2ºC/min. The FAME were identified by comparing their fragmentation pattern with authentic standards (Sigma) and also with NIST library (Dayananda et al. 2006). The acetone extract of the alga B. braunii (CFTRI-1) was analysed by HPLC using a reversed phase C18 column ( 25 cm x 4.6 mm ID) with an isocratic solvent system consisting of acetonitrile/methanol/dichloromethane (7:1:2) at a flow rate of 1.0 ml/min and the compounds were detected at 450 nm. Lutein, β-carotene were identified using authentic standards (Sigma). The cells were processed for scanning electron microscopy (SEM) according to Fowke et al. (1994). The samples were fixed in 2% glutaraldehyde in 0.2 M phosphate buffer (pH 6.8) for 12 hrs, dried in alcohol series up to 100%, sputter coated with gold and examined in a LEO Scanning Electron Microscope 435 VP (Leo Electron Microscopy Ltd.,Cambridge , UK ). ISSR fingerprinting of algal strains Genomic DNA was isolated using plant genomic DNA isolation kit (Sigma). Three inter simple sequence repeats (ISSR) primers (AC) 8C , (GA) 8C , (GA)8T were used for ISSR fingerprinting. The amplifications were carried out in 1X PCR buffer with 1.5 mM MgCl2, 2.5 mM of dNTPs and 1 unit of Taq DNA polymerase (MBI fermentas, Lithuania), 200 nM of each primer (Sigma). PCR was performed in a thermal cycler (Eppendorf Thermal Cycler) with a temperature programme involving initial denaturation at 94ºC for 5 min followed by 35 cycles of 30 sec denaturation at 94ºC , 1 min at respective annealing temperature and 1 min extension at 72ºC with a final extension of 72ºC for 10 min. The amplification products were separated by size on 1.2% agarose gels and stained with ethidium bromide observed under UV light and documented (Hero-Lab Gmbh. Germany). The PCR reactions were repeated at least twice to establish reproducibility. It has been reported that, Botryococcus braunii exists in the form of blooms in fresh water bodies like ponds, lakes and reservoirs (Metzger and Largeau, 2005). The samples collected in Bear Shola Falls (Kodaikanal , India ) were initially cultured in modifiedChu 13 medium and were purified by serial dilution fallowed by plating. The microscopic observations (Figure 1 and Figure 2) of the isolated alga revealed its colonial existence. Komárek and Marvan (1992) proposed the existence of at least 13 species of Botryococcus on the basis of morphological differences by omitting the chemical analyses. Metzger and Largeau (2005) reported that, within in each chemical race and for the same strain the morphology of the alga could vary in relation to age and culture conditions. The morphological heterogeneity of the alga makes the identification difficult. So in the present study we used ISSR as a tool to identify the alga by comparing with the known strain and also the nature of hydrocarbons it synthesize. ISSR finger printing (Figure 3) revealed a very close genomic similarity among the known B. braunii (B. braunii N-836, National Institute for Environmental Studies,Tsukuba , Japan ) and the isolated unknown alga. ISSR fingerprinting is a very potent tool to compare genomes for their identity. In some cases it is also possible to identify even minor somaclonal variations among the individuals (Rani et al. 2000; Li and Ge, 2001). However epigenetic variation related to change in DNA structure or methylation could not be explained by ISSR markers. ISSR markers have been successfully used in the determination of taxonomical relationships among species and subspecies (Hao et al. 2002; Vijayan et al. 2004). However, phylogenetic analyses of several strains of B. braunii were carried by 18s rRNA sequence (Senousy et al. 2004). Based on the 18s rRNA sequence analysis they showed that, the four strains of B. braunii belong to three existing chemical races which form a monophyletic group. The liquid culture developed from the single colonies was established in modifiedChu 13 liquid medium and the cultures were maintained by sub culturing at two weeks intervals. The growth curve (Figure 4) shows that the alga grew exponentially up to two weeks followed by stationary phase. It is also evident from Figure 5, that the decrease in phosphate concentration in the medium indicates the utilization of phosphate for its active growth. After two weeks of growth, phosphate and protein in the spent medium increased and correlate with the stationary phase. The increase in both phosphate and protein concentrations in the spent medium may be due to lyses of the cells. Similar observations by Casadevall et al. (1985) demonstrated the utilization of phosphate in the early exponential growth of the alga and increased amount of phosphate in the final stages of the culture. The early stationary phase culture was harvested, and the hydrocarbons were extracted from the dry biomass with hexane. Hydrocarbons were identified by their M+ ions and comparison of the mass spectra with those of the standards (Sigma) and also with the NIST library. The types of hydrocarbons produced by the alga were identified as saturated hydrocarbons in the range of C21 to C33 (peak 1 to peak 13) by GCMS (Table 1; Figure 6 and Figure 7). Among the saturated hydrocarbons produced by the alga, tetracosane (peak 4) and octacosane (peak 8) were found to be the major and constituted 17.6% and 14.8% respectively, and their mass spectra and gas chromatograms have been presented in Figure 6 and Figure 7. Yang et al. (2004) reported that the alga B. braunii (obtained from Culture Collection of Algae, Institute of Hydrobiology , Chinese Academy of Sciences) produced saturated hydrocarbons in the range of C21 to C31 and heptacosane as the major constituent. It has been also reported that the B. braunii produced saturated straight-chain and branched-chain (C14-C28) and long-chain linear aliphatic (C20-C27) hydrocarbons in the range of C12-C32 (Banerjee et al. 2002; Volova et al. 2003). Thus the ISSR analysis and hydrocarbon profile of the algal isolate clearly indicates that the Indian isolate is Botryococcus braunii.
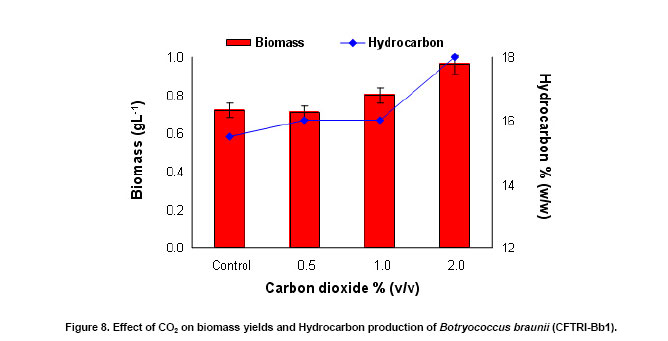
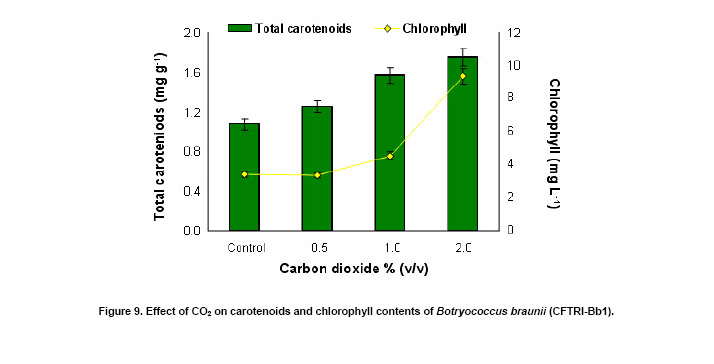
An increase of 1.1 to 1.3 fold in biomass yield was observed in B. braunii culture supplemented with carbon dioxide over the control (Figure 8). Similarly hydrocarbon accumulation also increased with the supplementation of carbon dioxide (Figure 8). Increase in chlorophyll and carotenoids contents was observed with increase in carbon dioxide supplementation up to 2% (v/v) (Figure 9). This shows its effective utilization of carbon dioxide through photosynthesis. Tripathi et al. (2001) reported photoautotrophic growth of different microalgae for higher growth and carotenoid production. It was evident that different algae require different levels of CO2 for their photoautotrophic adaptability (Tripathi et al. 2001). In the present study 2% of CO2 supplementation was found to be better for growth and hydrocarbon production. The fat content of the organism was found to be 22% (w/w) while palmitic and oleic acids as the major fatty acids constituting 40.6 and 22.3% respectively (Table 2). Similar observations were made by Fang et al. (2004) and Dayananda et al. (2006) and they reported that palmitic acid and oleic acids as the major components in the B. braunii.
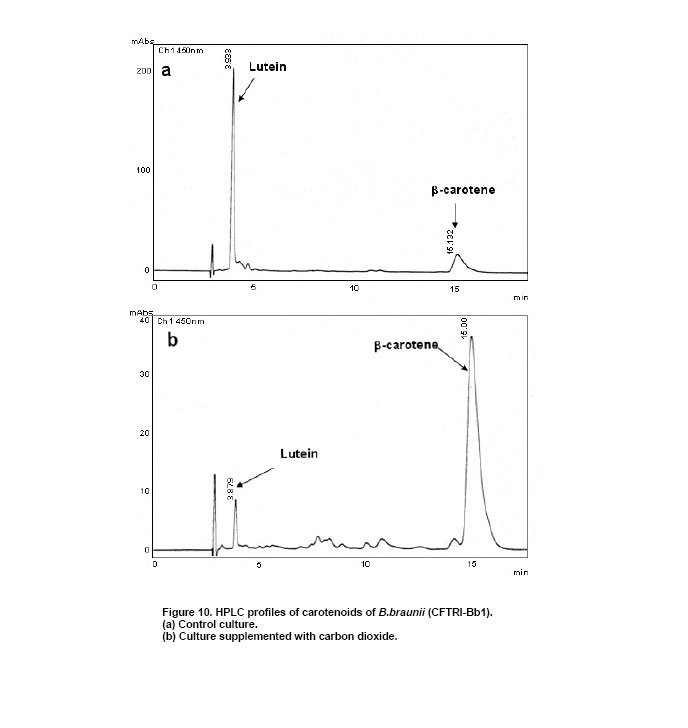
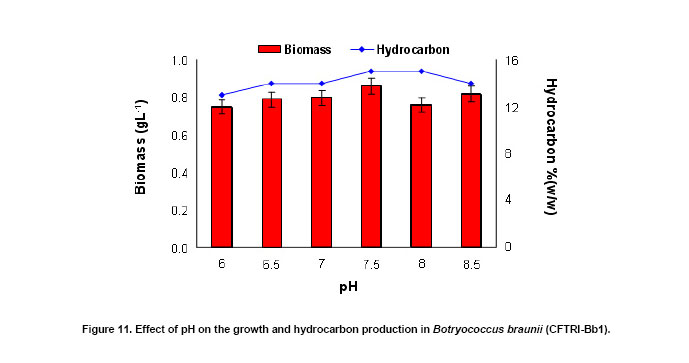
Lutein (64.1%) and β-carotene (25.1%) were found to be the major carotenoids (Figure 10) during the early stationary phase of the culture. In CO2 supplemented cultures, there was a 3.2 to 3.6 fold increase in β-carotene content (Figure 10) and total carotenoids content of the organism which enhanced (up to 1.75 mg g-1) with increasing carbon dioxide concentration (Figure 9). From Figure 11 it is evident that there is no significant effect of pH on the biomass yield and production of hydrocarbons since they varied in the range of 0.75 gL-1 to 0.86 gL-1 and 13 to 15% (w/w) respectively. However at pH 7.5 the algae showed maximum growth and production of hydrocarbons (Figure 11). In conclusion, the alga B. braunii (CFTRI-Bb1) can be of use in production of hydrocarbons. The supplementation of CO2 enhanced both biomass and hydrocarbon production. The organism exhibited wide range of pH adaptability. With further understanding on the influence of cultural conditions on hydrocarbon production, the alga can be exploited for outdoor cultivation. We thank Dr. V. Prakash, Director, CFTRI for his encouragement in carrying out this study and Dr. Narasimha Rao, Regional Director, Indira Gandhi National Open University,Bangalore for his need full help and advice during sample collection.
Note: Electronic Journal of Biotechnology is not responsible if on-line references
cited on manuscripts are not available any more after the date of publication.
© 2007 by Pontificia Universidad Católica de Valparaíso -- Chile The following images related to this document are available:Photo images[ej07008f10.jpg] [ej07008f7.jpg] [ej07008f6.jpg] [ej07008f2.jpg] [ej07008f11.jpg] [ej07008f8.jpg] [ej07008f5.jpg] [ej07008f9.jpg] [ej07008f3.jpg] [ej07008f4.jpg] [ej07008f1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}