 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Electronic Journal of Biotechnology, Vol. 10, No. 2, April 15, 2007, pg. 178-190 RESEARCH ARTICLE A practical approach to the understanding and teaching of RNA silencing in plants Ariel A. Bazzini1, Vanesa C. Mongelli2, H. Esteban Hopp3, Mariana del Vas4,Sebastián Asurmendi*5 1Instituto Nacional de Tecnología Agropecuaria
Instituto de Biotecnología
Las Cabañas y Los Reseros B1712WAA Castelar, Argentina
Tel: 5411 4621 1447. Ext 146
Fax: 5411 4621 0199
E-mail: abazzini@cnia.inta.gov.ar Financial support: This research was partially supported by grants PICT 11196, PID 267 and PAV 137 from FONCyT-ANPCyT and PIP 5788 from CONICET and other funding from INTA, Code Number: ej07018 Abstract Gene silencing, also called RNA interference (RNAi) is a specific mechanism of RNA degradation involved in gene regulation, development and defense in eukaryotic organisms. It became an important subject in the teaching programs of molecular biology, genetics and biotechnology courses in the last years. The aim of this work is to provide simple and inexpensive assays to understand and teach gene silencing using plants as model systems. The use of transient and permanent transgenic plants for expressing reporter genes, like those derived from jellyfish green fluorescent protein (gfp) encoding gene, provides a nice, colorful and conclusive image of gene silencing. Three experimental approaches to evidence RNA silencing are depicted. In the first approach gene silencing is demonstrated after transient expression of reporter genes in non-transgenic plants. In the second, silencing is triggered against a reporter gene stably integrated into a transgenic plant. The third approach involves the triggering of RNA silencing against endogenous genes using viral vectors. In addition we illustrate systemic gene silencing showing how the silencing signal is spread over a plant and finally it is also demonstrated the suppression of gene silencing. The first group of experiments is recommended to be tough on undergraduate courses, the following two sections are recommended for graduate courses. Hopefully, it will help students to understand this important phenomenon and to unravel the importance of gene silencing as a key gene regulation mechanism and as a molecular and biotechnological tool. Keywords: gene silencing, GFP, HC-Pro, PDS, protocols, PTGS, teaching, transgenic, transient expression, TRV, VIGS.
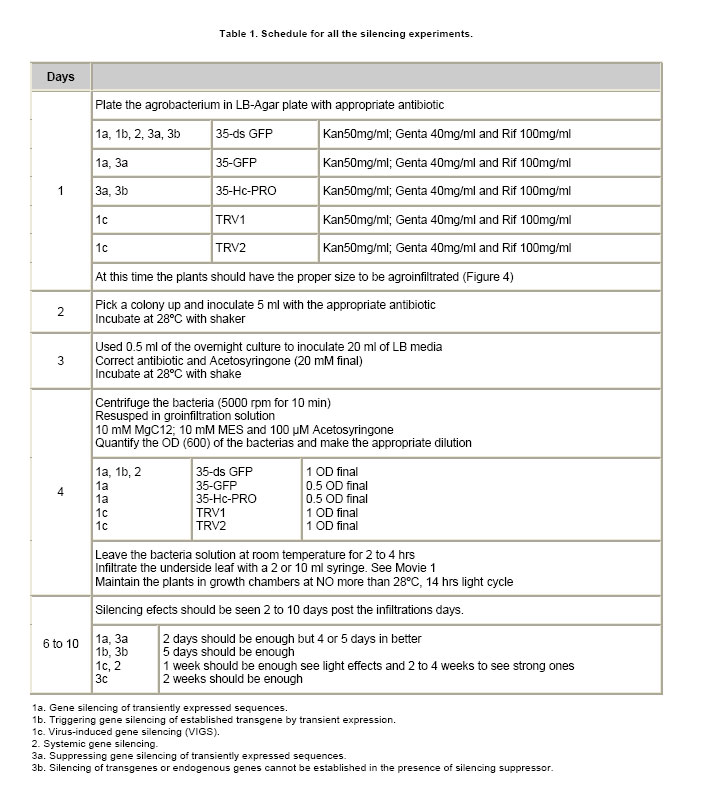
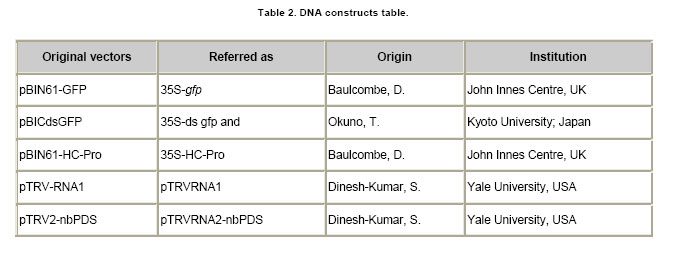
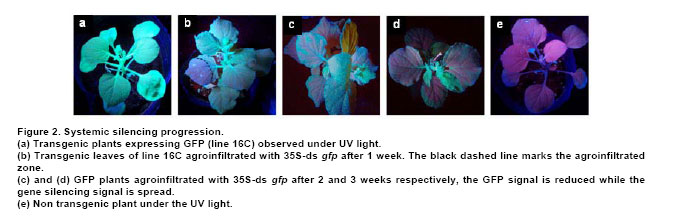
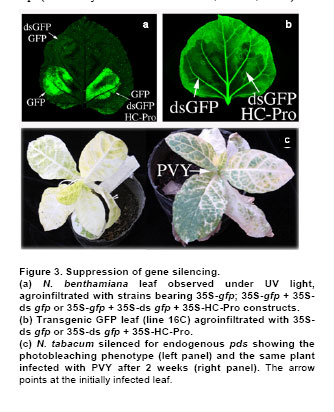
Post-transcriptional gene silencing (PTGS) in plants, known as RNA interference (RNAi) in animals, is an inducible and specific mechanism of RNA degradation (Baulcombe, 2004). It was shown that gene silencing is not only an important gene regulation mechanism but also a defense mechanism against foreign or aberrant endogenous RNAs, like virus and retrotransposons (Aravin et al. 2001; Hamilton et al. 2002). Due to the important role of RNA silencing per se and the multiple biotechnological uses that derived from the understanding of this phenomenon, we consider that this topic should be taught to biology students. We exemplify here a simple practical approach to the understanding and teaching of RNA silencing in plants. Molecular basis of RNA silencing Although RNA silencing operates through diverse pathways, it is most efficiently triggered by double stranded RNA (dsRNA) intermediates, which are processed into RNA duplexes of 21-24 nucleotides by a ribonuclease III-like enzyme called Dicer (Fire et al. 1998; Bernstein et al. 2001; Wesley et al. 2001). Once produced, these small RNA molecules or short interfering RNAs (siRNAs) are incorporated in a multi-subunit complex called RNA-induced silencing complex or RISC (Hammond et al. 2000; Tang et al. 2003). RISC is formed by a siRNA and an endonuclease, among other components. The siRNAs within RISC acts as a guide to target the degradation of complementary messenger RNAs (mRNAs) (Hammond et al. 2000; Tang et al. 2003). The summary of the silencing mechanism can be seen in Power Point Presentation 1. More recently, a closely related mechanism of RNAi was described for the regulation of the expression of endogenous genes including some involved in developmental processes. The host genome codify for small RNAs called microRNAs or miRNAs that are responsible of the endogenous gene silencing. The dsRNAs triggering of silencing can originate from several sources such as expression of endogenous or transgenic antisense sequences, expression of inverted repeated sequences or RNA synthesis during viral replication (Voinnet, 2005a). When dsRNA molecules produced during viral replication trigger gene silencing, the process is called virus-induced gene silencing (VIGS) (Lu et al. 2003). One interesting feature of RNA silencing in plants is that once is triggered in a certain cell, a mobile signal is produced and spread through the whole plant causing the entire plant to be silenced (Dunoyer et al. 2006). RNA silencing and viral infections It was proposed that being RNA silencing an antiviral defense mechanism, viruses must have evolved a counter-defense strategy (Mallory et al. 2001; Ratcliff et al. 2001). Consistent with this hypothesis, in the last 8 years several laboratories have discovered and characterized viral-encoded molecules with the capacity to suppress gene silencing (Brigneti et al. 1998; Anandalakshmi et al. 2000; Lucy et al. 2000; Zhou et al. 2006). Such molecules have also been found in a large number of plant and animal viruses (Anandalakshmi et al. 1998; Brigneti et al. 1998; Li et al. 2002). Nowadays, it is expected that practically most viruses carry suppressors of gene silencing, which are diverse in sequence, evolutionary origin, strength and mode of action (Roth et al. 2004). Among them, the helper component proteinase (HC-Pro) codified by Potyvirus genomes is a well known suppressor of gene silencing that prevents the accumulation of siRNAs and, as consequence, the establishment of gene silencing (Vance, 1991). Experimental approaches to demonstrate RNA gene silencing Even though gene silencing exists among eukariotic organisms, plants are the most widely used model system to illustrate gene silencing, because they are easy and inexpensive to handle (Fire et al. 1998; Voinnet 2005a). Transient expression assays in plants can be performed by a procedure called agro-infiltration that makes use of the bacterial plant pathogen Agrobacterium tumefaciens. During infection, Agrobacterium transfers T-DNA that is part of its plasmidial DNA to the infected plant. The Agrobacterium strains employed for infiltration contain the plasmid carrying the sequences in study, which will be expressed in the plant tissue. The technique consists in the infiltration of the abaxial surface of a leaf with an Agrobacterium suspension using a needle-less syringe (Movie 1). One well suited plant model to this approach is Nicotiana benthamiana, whose leafs can be easily infiltrated and produce high levels of protein. In order to perform gene silencing assays in plants, reporter genes are used that allow a rapid and quantitative way to distinguish if they are active or silenced. The jellyfish gene encoding for the green fluorescent protein (gfp) is one of the most widely used reporter genes. Its expression results in a green fluorescent emission under UV light (Stearns, 1995). Due to chlorophyll emission, wild type healthy plant tissues look bright red under UV light; therefore areas expressing GFP contrast with non agroinfiltrated areas exposed to the same light source. Other frequently reporter genes used in gene silencing assays are those implicated in the biosynthesis of plant pigments. Silencing induction of this type of genes leads to the reduction of colored compounds accumulation, a phenomenon that can be easily noticed with just bare eyes (Fofana et al. 2004). For example, phytoene desaturase protein (PDS) is a key enzyme in the carotenoid biosynthesis pathway. Silencing of the PDS gene (pds) results in white leaves, a phenotype called photobleaching. This work is focused to: 1. Triggering of gene silencing; 2. Systemic gene silencing and finally 3. Gene silencing suppression. The first topic, describes experiments that illustrate the phenomenon of gene silencing of transgenes or endogenous genes, triggered by either transient expression of homologous sequences or viruses carrying them. This section is recommended to be taught in basic courses such as Genetics and Molecular Biology. The second topic illustrates systemic gene silencing showing how the silencing signal is spread over a plant. Finally the third topic exemplifies suppression of gene silencing. The second and third sections are orientated to more advanced courses in Biotechnology and Virology areas. These experiments were recently used as experimental practice for a Molecular Phytopathology course taught at the University of Buenos Aires, Argentina. The aim of this work is to provide simple and inexpensive assays to teach gene silencing using plants as model systems. These experiments provide a nice, colorful and conclusive image of plant gene silencing. Hopefully, it will help students to understand this important phenomenon and to unravel the importance of gene silencing (and its suppression) as a key gene regulation mechanism and as a molecular and biotechnological tool. Materials and MethodsTransgenic and non-transgenic plants were grown under standard greenhouse conditions or maintained in growth chambers at 24 to Agrobacterium infiltration procedure Agrobacterium tumefaciens strain GV3101 carrying the different constructs were plated in LB-Agar medium with an appropriate antibiotic (Table 1) and maintained at The following DNA constructs were used: 35S-gfp, 35S-ds gfp and 35S-HC-Pro (Dr. David Baulcome from the John Innes Centre, UK) and TRV constructs pTRVRNA1 and pTRVRNA2-nbPDS (Professor Savithramma Dinesh-Kumar from Yale University, USA), Table 2. Plants expressing GFP were observed under a strong UV lamp (Black ray B-100AP 100 Watts, UVP®, USA) in a completely dark room. To use the UV lamp it is required to wear UV protection goggles and avoid exposure of the naked skin to the UV light. Whole plants pictures for this work were taken using a Kodak digital camera using a UV filter. The GFP florescence of the agroinfiltrated leaves for this work was documented using a Typhoon Scanner (Amersham Biosciences, UK). ResultsHere we describe three experimental approaches to evidence RNA silencing. In the first approach gene silencing is demonstrated after transient expression of reporter genes in non-transgenic plants. In the second, silencing is triggered against a reporter gene stably integrated into a transgenic plant. The third and last approach involves the triggering of RNA silencing against endogenous genes using viral vectors. Gene silencing of transiently expressed sequences. Infiltration of non-transgenic Nicotiana benthamiana leaves with a strain of Agrobacterium tumefaciens carrying a gfp gene construct (35S-gfp) results in transient GFP expression, observed as a green fluorescence under UV illumination (Figure 1a) (Llave et al. 2000; Voinnet et al. 2000; Dunoyer et al. 2002; Hamilton et al. 2002; Takeda et al. 2002; Bucher et al. 2003). In contrast, co-infiltration of agrobacteria carrying a 35S-gfp construct together with a strain carrying an inverted repeat of gfp construct (35S-ds gfp) does not produce any green fluorescence. This phenomenon is a consequence of gene silencing triggered by 35-ds gfp resulting in a drastic reduction of the GFP level (Figure 1a). In this experiment, siRNAs are produced from the 35S-ds gfp construct; those siRNAs (homologous to the gfp sequence) guide the recognition by RISC of GFP mRNAs expressed from the 35S-gfp construct that in turn leads to its degradation (Slide 2 Power Point Presentation 1). Therefore, no fluorescent signal is detected and the plants look red due to normal chlorophyll emission, the same as mock-infiltrated controls under UV light. Triggering gene silencing of stable transgene by transient expression. It is also possible to trigger PTGS of endogenous genes or stable transgenes through transient expression of homologous sequences. A N. benthamiana transgenic line expressing GFP (line Virus-induced gene silencing (VIGS). The most commonly used viruses to induce RNA silencing are Tobacco rattle virus (TRV) and Potato virus X (PVX) (Lu et al. 2003; Burch-Smith et al. 2004; Robertson, 2004). Both produce mild symptoms in Solanaceae plants species that do not interfere with the outcome phenotype. The infection is carried out by agro-infiltrating plant tissue with a plasmid vector containing an infective copy of the viral genome. The viral genomes are genetically modified to allow the expression of a foreign sequence from a viral promoter. PVX and TRV have positive single stranded RNA genomes, therefore the transcription of one copy of the viral genome from the plasmid vector results in the generation of one infective virus that can perfectly replicate and infect the whole plant. TRV has a bipartite genome formed by RNA1 and RNA2 molecules. RNA1 contains genes essential for virus replication, including the replicase and movement protein. It can replicate and move systemically without the presence of RNA2. RNA2 codes for the coat protein and for two non-structural proteins that are required for virus spread from plant to plant. Liu et al. (2002) engineered two constructs harboring the RNA1 or RNA2 under the 35S promoter called TRV1 and TRV2 respectively (Wesley et al. 2001). TRV2 was further engineered and the two non structural genes were removed and a multiple cloning site was added; therefore any sequence can be easily cloned into the virus genome and then expressed together with the virus replication (Ratcliff et al. 2001). Inoculation of N. benthamiana plants with a modified TRV (TRV1 and TRV2) carrying host-derived inserts, produces a reduction of the expression of the endogenous homologous gene 7 to 15 days post infection. Figure 1c, shows N. benthamiana and N. tabacum plants infected by means of agro-infiltration using the infectious TRV constructs, carrying a modified TRV2, harboring a portion of 400 bp sequence from pds gene show a photobleaching phenotype The leafs look almost completely white due to the silencing of the endogenous gene (photobleaching phenotype). As a conclusion for this section, gene silencing is a natural phenomenon in plants that is triggered by dsRNAs. This dsRNA molecules can be originated by the expression of constructs bearing sense and antisense sequences (which transcription give rise to dsRNAs) and by viral replication. Any of these approaches can be used to study the effect of the functional knock out (gene silencing) of any desired gene. As mentioned in the introduction, the silencing signal spreads over the entire plant. Here we illustrate the spread of the gene silencing signal from the agroinfiltrated zone to the entire plant giving rise to systemic gene silencing. When a lower leaf of a N.b. As a conclusion, gene silencing initiates where it is triggered, then spreads along the entire leaf and finally moves from one leaf to the others using the vascular tissues to ultimately reach the entire plant. It was demonstrated that the cell-to-cell mobile silencing signal is a 21 nt siRNA molecule that is spread using a “relay” process where it is re-amplified by a cellular RNA-dependent RNA polymerase every 10-15 cells (Himber et al. 2003; Yoo et al. 2004; Voinnet, 2005b). As mentioned before, viruses have evolved to encode for proteins that suppress gene silencing. In this section we will exemplify gene silencing suppression using the same experimental approaches as before but adding the expression of the well characterized potyviral suppressor, HC-Pro. Finally, we will show how a virus can suppress an established gene silencing during a natural infection process. Suppressing gene silencing of transiently expressed sequences. As shown in Figure 1a, transient expression of 35S-gfp gives rise to a bright green area in a non-transgenic plant under the UV light (Figure 3, lower left side). In contrast, when 35S-gfp and 35S-ds gfp are co-expressed, no florescence is detected because of GFP silencing triggered by the ds gfp construct (Figure 1a and Figure 3a, upper right side). When a strong gene silencing suppressor as HC-Pro, is co-expressed with constructs 35S-gfp and 35S-ds gfp, the infiltrated zone remains bright green (Figure 3a, lower right side of the leaf). This is due to the inhibition of the silencing machinery caused by HC-Pro. Specifically HC-Pro inhibits an intermediate step of RNA silencing via binding to siRNAs. In addition it inhibits the siRNA-initiated RISC assembly pathway by preventing RNA silencing initiator complex formation. (Lakatos et al. 2006). Silencing of transgenes or endogenous genes cannot be established in the presence of a silencing suppressor. Gene silencing suppressors can also inhibit silencing of transgenes or endogenous genes. As shown in Figure 1b, when leaves of the N.b. Suppression of gene silencing mediated by virus infection. Gene silencing can be permanently triggered in transgenic plants by designing and using a transgene containing a sense and antisense copy of the gene to be silenced in such a way that a dsRNA is produced (Wesley et al. 2001). Nicotiana tabacum and Nicotiana benthamiana silenced for the magnesium chelatase gene show a characteristic photobleaching phenotype (Figure 3c, left plant) (Fofana et al. 2004). However, the photobleaching phenotype is markedly reduced and the green pigmentation restored (Figure 3c, right plant) when the silenced plants are infected with PVY. This indicates that the replication of PVY was able to suppress the established magnesium chelatase silencing. As we previously mentioned PVY carries a gene encoding for a strong silencing suppressor called HC-Pro. Therefore, proteins able to suppress gene silencing can be easily detected through this experimental system and so any viruses that may carry them. Most of the viruses have some sort of silencing suppressor system as an evolutionary counter-response to the antiviral activity of the plant gene silencing defense. Concluding RemarksRNA silencing has become a hot topic of biological research in the last few years. Understanding gene silencing is highly relevant to comprehend the interaction between virus and their hosts and, in a wider view, to comprehend the relationships between foreign RNAs (including retrotransposons, viroids, etc) and eukaryotic cells. Moreover, gene silencing reveals an entirely new level of post-transcriptional gene regulation. Furthermore it became an extremely useful technique for molecular biology and a very powerful biotechnological tool. Here, we performed a detailed description of the most common assays related to gene silencing that will provide simple and inexpensive tools to help to the understanding and teaching of RNA silencing. We consider the topic is of vital importance for the students interested in biotechnology and plant pathology. AcknowledgmentsWe are grateful to Dr. David Baulcombe/Sainsbury Laboratory/The Gatsby Charitable Foundation. (John Innes Centre) for the GFP
Note: Electronic Journal of Biotechnology is not responsible if on-line references cited on manuscripts are not available any more after the date of publication. © 2007 by Pontificia Universidad Católica de Valparaíso -- Chile The following images related to this document are available:Photo images[ej07018t1.jpg] [ej07018f8a.jpg] [ej07018f6a.jpg] [ej07018f6c.jpg] [ej07018f6b.jpg] [ej07018f7.jpg] [ej07018f4.jpg] [ej07018f1.jpg] [ej07018f2.jpg] [ej07018t2.jpg] [ej07018f5.jpg] [ej07018f8c.jpg] [ej07018f8b.jpg] [ej07018f3.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}