 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
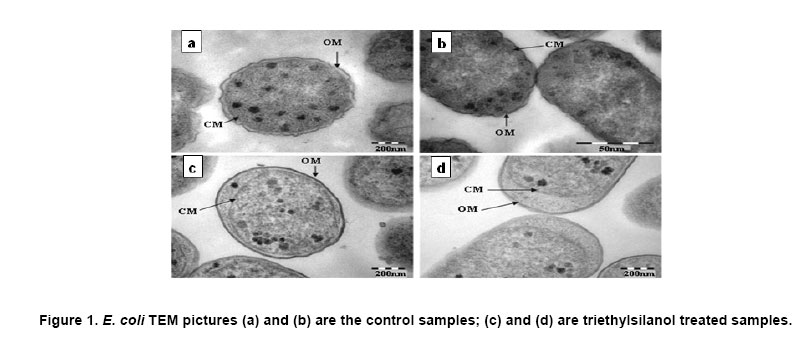
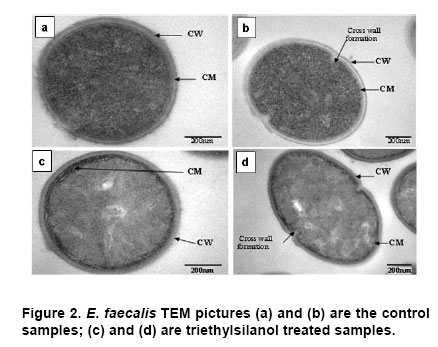
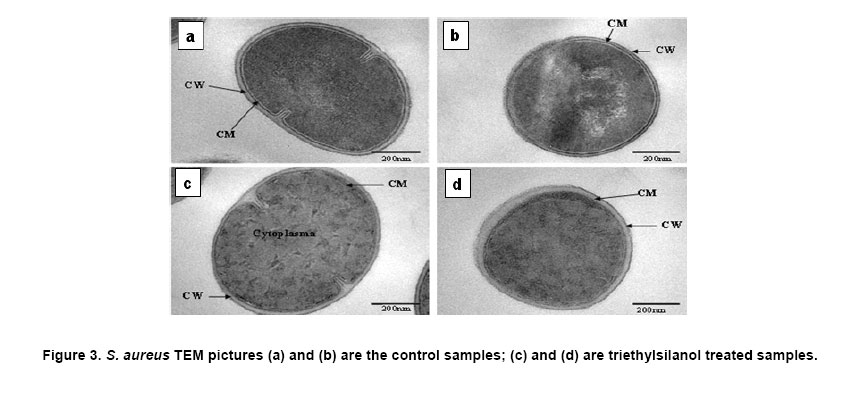
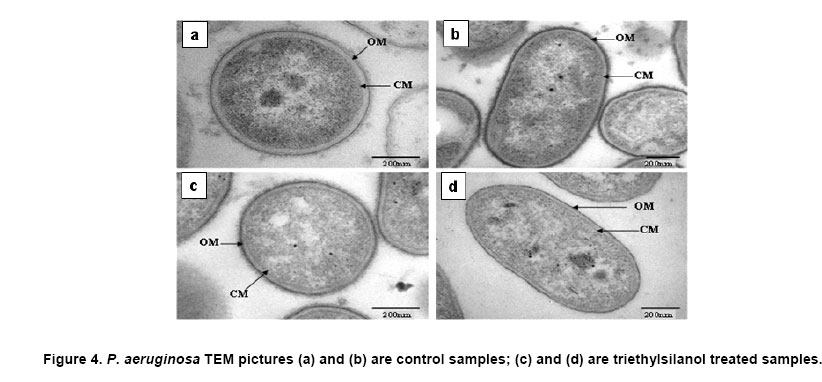
Electronic Journal of Biotechnology, Vol. 10, No. 2, April 15, 2007, pg. 252-259 RESEARCH ARTICLE Membrane damage of bacteria by silanols treatment Yun-mi Kim1, Samuel Farrah2, Ronald H. Baney*3 1 Department of Materials Science and Engineering
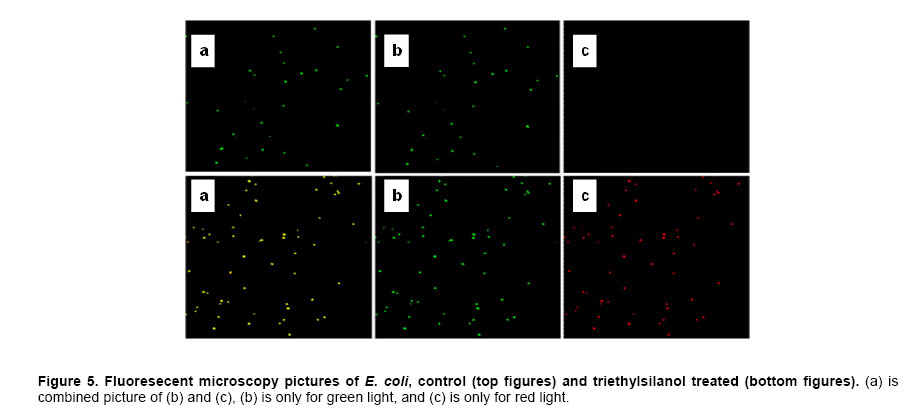
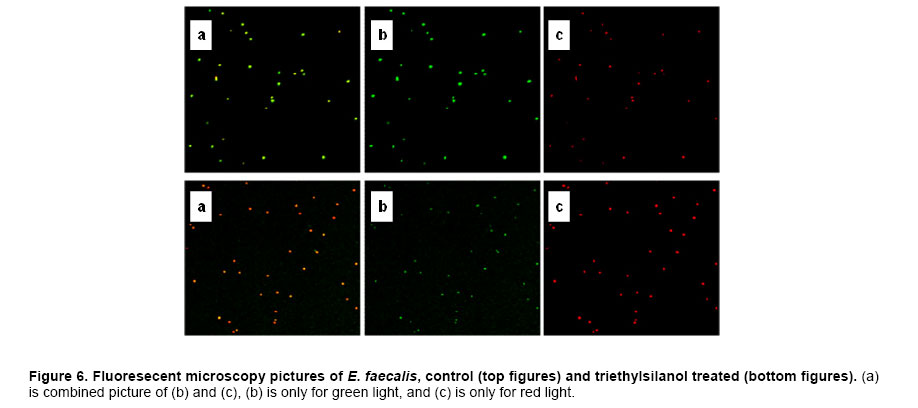
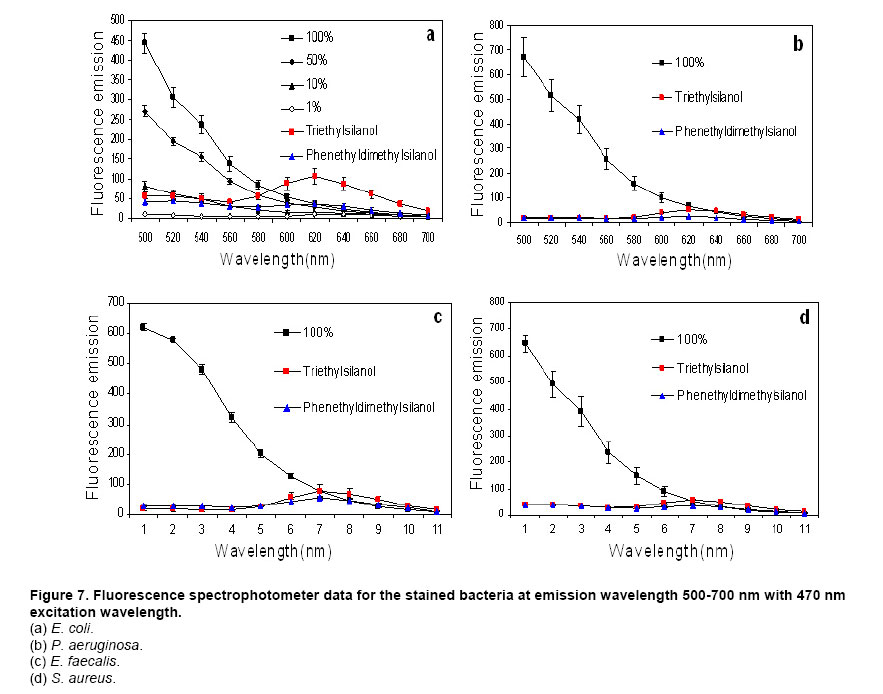
Website : http://baney.mse.ufl.edu/ Financial support: Air Force Research Lab. in Tyndall Air Force Base. Code Number: ej07025 Abstract Antimicrobial action of silanols, a new class of antimicrobials, was investigated by transmission electron microscopy and fluorescent dye studies. Gram-negative bacteria, Escherichia coli and Pseudomonas aeruginosa and Gram-positive bacteria, Staphylococcus aureus and Enterococcus faecalis were treated by silanols at concentration of less than 0.2 wt% for one hour. Membrane damage of the bacteria by the silanol treatment was clearly observed by transmission electron microscopy. Separation of the cytoplasmic membrane from the outer membrane for E. coli and disorganized cytoplasmic membrane of the Gram-positive bacteria were observed when compared to the control. Fluorescent dyes, green-fluorescent nucleic acid stain (Syto 9) and the red-fluorescent nucleic acid stain (Propidium iodide), were used to monitor membrane damage of the bacteria by Confocal microscopy and Spectrophotometer. A reduction of the green fluorescent emission was detected for silanol treated bacteria indicating membrane damage of the bacteria and supporting the hypothesis that their viability loss may be due to their membrane damage analogus to alcohols. Keywords: antimicrobials, hydrophobicity, organosilicon compounds. Silanols, alcohols, and aromatic alcohols (phenols) have similar chemical structures consisting of a hydrophilic hydroxyl group and hydrophobic organic substitutents. Alcohols and phenols are widely used antimicrobial agents while the silanols were recently discovered to be strong biocidal agents in comparison with structurally analogous carbinols (Kim et al. 2006). Silicon is more electropositive than the carbon. The silicon atom is larger than the carbon atom and it can accommodate extra coordination providing. For example, the enzymatic action of the recently reported silicatine is believed to proceed through extra coordination at silicon (Shimizu et al. 1998). Both effects could provide possible different interaction pathways with cells. It is not unreasonable to suggest, however, that the antimicrobial mode of actions of the silanols might resemble those of the alcohols and the phenols. It is generally known that alcohols cause membrane damage (Mcdonnell and Russell, 1999; Etschmann et al. 2002) through an increase of the lipid solubility. In other words, alcohols increase the membrane fluidity through disorder of the biological self-assembly of the membrane. The primary target site of the phenolic compounds is the cytoplasmic membrane of bacteria (Mcdonnell and Russell, 1999). The damage to the cytoplasmic membrane causes the membrane to lose its ability to be a permeability barrier, subsequently leading to a loss of structural integrity and a leakage of intracellular material. Silanols are environmentally friendly materials because they are rather quickly degraded into environmentally benign silica, carbon dioxide and water in the environment (Graiver et al. 2003). Silanols can be applied by a various delivery systems such as a neat liquid phase, vapor phase due to their high volatility. In addition, unlike their analgous carbinols, silanols can be employed as controlled release agents through release by hydrolysis of Si-O-C bonds from carbinol baring substrates. This is quite analogus to the “blocking agents” employed in the pharmaceutical industry. These novel biocide silanols can be prepared from low cost intermediates derived from the commercial processes associated with the silicone industry. A bacterium has several layers of material that enclose or protect its cytoplasm (Moat et al. 2002; Barton, 2005). The cytoplasm region contains critical life units protected by the plasma membrane and the cell wall. In the Gram-positive bacteria, such as S. aureus and E. faecalis, the cell wall is thick (15-80 nanometers) consisting of several layers of peptidoglycan. On the other hand, Gram-negative bacteria such as E. coli and P. aeruginosa have a relatively thin (10 nanometers) cell wall surrounded by an outer membrane. The membranes are a bilayered structure and a selective permeability barrier that regulates the passage of substances into and out of the cell. The plasma membrane is the last barrier that can separate or protect the cytoplasm from the environment (Moat et al. 2002; Barton, 2005). Both electrostatic and hydrophobic interactions are involved in maintaining the spatial organization of the membrane components (Ingram and Buttke, 1984). It is well accepted that disorganization of the membrane by undesired or foreign substances can cause loss of the permeability or of the integrity of the membrane. This ultimately cause the death of the cell. For many antimicrobial agents, antimicrobial actions are initiated by interactions of the biocides with the cell wall membrane of the microorganisms. The agents then penetrate into the cell and finally act at the target sites (Mcdonnell and Russell, 1999). Hunt (1975) proposed that the potency of aliphatic alcohols is directly related to their lipid solubility. The lipid in the membrane bilayers can be dissolved by alcohols through the induced hydrophobic interaction between the alkyl chain from the alcohols and the lipid region in the membrane (Hunt, 1975; Ingram and Buttke, 1984). The hydrophobicity is an important parameter with respect to toxicity or alternation of the membrane’s integrity because it is directly related to the membrane permeation, which can result in disruption of the membranes of the bacterium. A plasma membrane integrity test with short-chain aliphatic alcohols illustrated that an increase of the alkyl chain length from methanol to 1-octanol enhanced the loss of the membrane integrity of epithelial cells in vitro (McKarns et al. 1997). Those authors suggested that alteration of the membrane integrity correlates with the hydrophobic properties of the alcohols. In this study we suggest that the hydrophobic interaction occurs between the organic group of the silanols and the lipophilic portion of the membranes as well as the hydrogen bonding between the hydroxyl function of the silanol and either the ester linkages of fatty-acyl residues or water molecules (Ingram and Buttke, 1984). We hypothesize that the silanol can partition effectively into the membrane through these interactions and cause its disorganization by increasing lipid solubility. Transmission electron microscopy and fluorescent dye techniques were utilized to verify our hypothesis of the membrane damage on the bacteria treated by silanols. Materials and MethodsBacterial preparations and antimicrobial test procedure The bacterial strains employed were the Gram-negative bacteria, E. coli C3000 (ATCC 15597) and a laboratory strain of S. aureus provided by the Department of Microbiology at the University of Florida and the Gram-positive bacteria, P. aeruginosa type strain (ATCC 10145) and E. faecalis type strain (ATCC 19433). The bacteria suspensions were prepared according to the procedure by Rincón and Pulgarin (2003). The bacteria were inoculated into a flask containing 90 ml of autoclaved water and 10 ml of Columbia broth and incubated for 20-24 hrs at The antimicrobial activity tests were carried out in aqueous condition (Cremieux and Davin-Regli, 2000) by adding a given concentration of antimicrobial agent to 9 ml of medium, deionized water, and 1 ml of the bacterial suspension. Triethylsilanol received from Gelest, Inc. and synthesized phenethyldimethylsilanol were used as the antimicrobials. The purity of the silanols measured by H-1 NMR (Nuclear Magnetic Resonance) showed 95 ± 3% for triethylsilanol and 90 ± 3% for phenethyldimethylsilanol. The impurity of the silanols was disiloxane, a condensation by-product of the silanols. The antimicrobial activity of the disiloxane was insignificant (Kim et al. 2006). The solution was mixed for an hour with constant stirring at room temperature. One millilitre sample was collected after a 1 hour treatment for the viability test. Viabilities of treated bacteria were confirmed by the plate-count method (Harrigan, 1998; Spencer and Ragout de Spencer, 2001). TEM sample preparation procedures Bacterial suspensions, E. coli, P. aeruginosa, S. aureus, and E. faecalis, were treated by triethylsilanol at a concentration of 0.2 (% g/g) for 1 hr. The control was prepared from the same batch of bacterial suspension at the same condition as the treated one. The plate count method confirmed no bacteria growth after the triethylsilanol treatment. The control and the treated bacteria suspension were centrifuged at 2600 rpm for 5 min and washed with a buffer solution Fluorescent dye, “LIVE/DEAD” BacLight Bacterial Viability Kits provided by Molecular Probes, have the capability of monitoring the viability of the bacteria as a function of the cell’s membrane integrity (Bunthof et al. 2001; Laflamme et al. 2004). Bacteria with a compromised membrane that are considered to be dead or dying will stain red, whereas cells with an intact membrane will stain green. The Viability Kit (L7007) is composed of green-fluorescent nucleic acid stain (Syto 9) and the red-fluorescent nucleic acid stain (Propidium iodide). The red and the green stain show a difference in spectral characteristics, different emission wavelength, and in their ability to penetrate healthy bacterial cells. The green stain can label bacteria with intact membranes and with damage membranes. In contrast, the red stain penetrates only bacteria with damaged membranes and causes a reduction in the green fluorescence when both dyes are present. The excitation /emission range of the green stain is 470/510-540 nm and 470/620-650 nm for the red stain. The fluorescent dye kit has the green stain, Five microlitre of the stained bacterial suspension was placed between a slide and a cover slip for microscopy evaluation. MRC-1024 confocal laser scanning microscopy at the McKnight Brain Institute at the Fluorescence spectrophotometer One millilitre of the stained bacterial suspension was placed in a quartz rectangular spectrophotometer cell. The cell has Suprasil quartz windows with a Membrane damage of bacteria by silanols treatment - TEM Transmission electron microscopy revealed structural membrane damage for the four bacteria, E. coli, P. aeruginosa, S. aureus, and E. faecalis, treated by triethylsilanol for 1 hr at the minimum lethal concentration of 0.2 (% g/g). The minimum lethal concentration (MLC) was defined as the concentration required for a 7-log reduction in viable bacteria after 1 hr exposure period in this study. The plate count method supported a complete deactivation of the bacteria after the silanol treatment, meaning 99.99999% elimination of the viable bacteria. A smooth continuous membrane structure was clearly observed in the untreated bacteria while the treated bacteria showed relatively disorganized membrane structures. Primary fixation allows the membrane structures to be preserved. Glutaraldehyde cross linked the proteins and OsO4 cross linked the lipids. Since the triethylsilanol treated bacteria and the control cell were processed by the same procedure the differences observed were caused by the silanol treatment. The membranes were clearly identified in the TEM pictures due to the primary fixation of OsO4 and the post staining of the bacteria. The dark areas show where the sample had a high electron density and the light areas show where the sample had a low electron density (Anderson et al. 2004). A general observation of E. coli treated by triethylsilanol was that the cytoplasmic membrane (CM) appeared to be retracted from the outer membrane ( Membrane damage of the bacteria by silanols treatment - Fluorescent dye studies Membrane damage due to the silanols treatment was monitored by using fluorescent dye through fluorescence microscopy and fluorescence spectrophotometry. Four bacterial suspensions without the silanol treatment were tested as control and compared with bacteria treated by the silanols. As expected, the silanol treated bacteria exhibited a reduced green fluorescence light and an increased red fluorescence light. Penetration of the red dye into the bacteria with damaged membrane led to a reduction of the green light. As a result, the silanol treated bacteria showed a double staining fluorescence light with a strong red and a weak green light under fluorescence microscopy as displayed in the bottom figures of Figure 5 for E. coli and Figure 6 for E. faecalis. In comparison, bacteria without the silanol treatment showed mostly green light in the case of the Gram-negative bacteria as seen in the top figures of Figure 5, while a strong green light and a weak red light was detected for the Gram-positive bacteria as seen in the top figures of Figure 6. P. aeruginosa and S. aureus displayed the similar results. Three fluorescence microscopy pictures for each bacterium are displayed to verify the reduction of the green light in bacteria. In Figure 5, E. coli, figure A is a combined picture of B and C, B is only for green light and C is only for red light. It should be mentioned that bacteria showed in figure B and figure C from Figure 5 are exactly the same E. coli, indicating the double staining on the bacteria. E. faecalis in Figure 6 showed the similar results. Laflamme et al. (2004) characterized bacterial viability as a function of membrane integrity using fluorescent dye, SYTO 9 and Propidium iodide. In spores with compromised membrane, double staining was detected, and showed that the red light was brighter than the green light. Fluorescence spectrophotometry provides quantitative results (Wu et al. 1999) suggesting a reasonable explanation regarding the fluorescence microscopy results. Different concentrations of bacterial suspensions were prepared by diluting from 100% to 50%, 10%, and 1% as controls. The fluorescence emissions of each concentration were measured by the spectrophotometer at a wavelength of excitation of 470 nm and an emission of 500-700 nm. A relative intensity of the fluorescence emissions at the range between 500 and 550 nm in wavelength where the green light is detected was differentiated depending on the concentration of the bacteria as seen in Figure 7a. Fluorescence emission corresponding to 10% bacterial concentration represents a 1 log-reduction and 1% is for a 2 log-reduction of viable bacteria. Fluorescence emissions of bacterial suspensions treated by the silanols were compared with the controls. A significant reduction of fluorescence emission at the green light emission wavelength of 500-550 nm was observed for the treated bacteria when compared to the 100% control sample as shown in Figure 7. The red dye penetrated into the bacteria with compromised membrane and reduced the green light. An increase of red light emission at 620-650 nm wavelength was also displayed for the treated bacteria. The extent of increase of red light was relatively small except for E. coli with triethylsilanol treated shown in Figure 7a. The fluorescence spectrophotometer results showed only a 1 or 2-log reduction of viable bacteria for the silanol treated samples even though the plate count method confirmed a 7-log-reduction of the treated bacteria. This result is consistent with the fluorescence microscopy result showing double staining for treated bacterial suspension. In other words, the fluorescence emission did not show a zero emission of green light or indicate a 7-log-reduction because the green dye is present with the red dye as double staining for the treated bacteria. The reduction of the green fluorescence light as well as the increase of the red fluorescence light was observed as an indication of the membrane damage of the silanol treated bacteria. In summary, the mechanism of action of the silanols upon the cell membrane of the bacteria was investigated. A change in the membrane after the silanols treatment was detected by transmission electron microscope, showing a detachment of the plasma membrane from the outer membrane for E. coli. In the case of the Gram-positive bacteria a disorganized cytoplasmic membrane was observed upon the silanol treatment when compared to the smooth and intact cell membrane of the untreated bacteria. The fluorescence method allowed us to monitor bacteria viability based on the membrane damage because the red dye only penetrates into the bacteria with membrane damage, subsequently reducing the green fluorescence. A significant reduction of the green light, was demonstrated by fluorescent microscopy and spectrophotometer, suggesting that the bacteria treated by the silanols had compromised membranes. The membrane damage might be one of causes for the death of the viable bacteria. AcknowledgmentsWe are grateful to Dr. Ben Koopman for allowing us to use his Microbiology laboratory. The authors would like to thank to ICBR Electron Microscope Core Laboratory for TEM and McKinght Brain Institute for Confocal Microscope. References
Note: Electronic Journal of Biotechnology is not responsible if on-line references cited on manuscripts are not available any more after the date of publication. © 2007 by Pontificia Universidad Católica de Valparaíso -- Chile The following images related to this document are available:Photo images[ej07025f2.jpg] [ej07025f6.jpg] [ej07025f5.jpg] [ej07025f4.jpg] [ej07025f1.jpg] [ej07025f7.jpg] [ej07025f3.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}