 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Electronic Journal of Biotechnology, Vol. 10, No. 2, April 15, 2007, pg. 328 - 335 TECHNICAL NOTE Single embryoid body formation in a multi-well plate Uthayashanker R. Ezekiel*1 , Mariappan Muthuchamy2, Jan S. Ryerse3 , Rita M. Heuertz4 1 GeneProTech, Inc. *Corresponding author Code Number: ej07031 Abstract Embryoid bodies (EB) formed from murine embryonic stem (ES) cells recapitulate many aspects of a developing embryo. Of specific importance, synchronous differentiation of EB recapitulates organ-specific development and is achieved in culture by formation of uniformly sized EB. The method described here demonstrates a simple and cost-effective way of generating EB from murine ES cells. Single EB are formed in a multi-well plate format and large numbers of EB are generated using a 96-well multi-well plate. Uniform single-sized EB formed in the multi-well are an ideal system for screening compounds and determining differentiation effects. Since EB contain all three germ layers, they are appropriate for studying small molecule effects on differentiation of ES such as is performed in high-throughput screening protocols. Keywords: differentiation, EB, ES, murine embryonic stem cell.
Murine embryonic stem (ES) cells are pluripotent and derived from the inner cell mass of an early embryo. When cultured in the presence of anti-differentiation agents such as leukemia inhibitory factor (LIF) and embryonic fibroblasts, these cells maintain their pluripotency and have the ability to differentiate into any cell type of the body (Evans and Kaufman, 1981; Bain et al. 1995; Smith, 2001; Chen et al. 2003). Removal of the anti-differentiation agent causes the ES cell to spontaneously differentiate which tends to follow a reproducible temporal pattern and which, in many ways, recapitulates early embryogenesis. When ES cells are cultured in suspension without anti-differentiation agents, they form three-dimensional aggregates called embryoid bodies (EB). Within 2-4 days in suspension culture, endoderm form from the inner cell mass thereby giving rise to simple EB (Leahy et al. 1999). On approximately day 4, differentiation of columnar epithelium with a basal lamina and formation of a central cavity occurs. At this stage, the EB are called cystic embryoid bodies and bear similarity to the egg-cylinder stage of mouse embryos (Evans and Kaufman, 1981; Keller et al. 1993; Ng et al. 2005). Upon continued in vitro culture, EB give rise to all three germ layers (ectoderm, mesoderm, endoderm) and terminally differentiate into a wide variety of cell types, such as cardiomyocytes, hematopoietic cells, neurons and pancreatic islet cells (Risau et al. 1988; Bain et al. 1995; Smith, 2001; Dang et al. 2002; Liersch et al. 2006). From the time of implantation and throughout gastrulation, early mammalian development is characterized by rapid cellular proliferation and migration. In mouse embryogenesis, the transitory movements of gastrulation begin shortly after implantation (6.5 days post coitum [d.p.c.]) thereby placing the endoderm, ectoderm and mesoderm in their respective positions for organogenesis. Comparison studies of murine EB developmental patterns versus murine embryonic development have been performed using gene expression and tagged reporter constructs (Xiong et al. 1998; Leahy et al. 1999; Vallier et al. 2001). Results indicate that prior to day 3, EB in suspension culture are equivalent developmentally to pregastrulation-stage embryos (4.5 - 6.5 d.p.c). Between days 3 and 5, EB contain cell types present in embryos during gastrulation (6.5 - 7.5 d.p.c.). After day 6, EB are equivalent to embryos in the stage of early organogenesis (7.5 d.p.c). Differentiation of EB into particular cell lineages is an area of intensive study. Most of the methods utilized for EB differentiation studies involve several steps: 1) production of EB from suspension culture; 2) exposure of EB to agents designed to induce differentiation of a specific lineage; and 3) growth of EB on tissue culture plates coated with biological molecules such as gelatin or laminin (Snodgrass et al. 1992; Bain et al. 1995; Boheler et al. 2002; Dang et al. 2004). Since EB give rise to all types of cells and recapitulate events similar to those that occur during embryonic development, the use of EB to study effects of small molecules and/or biological agents is an ideal system. Indeed, there are many advantages to using EB as a model system instead of embryos or whole animals. For example, a large number of EB can be produced and used for throughput screening assays in a manner that is cost-effective, especially when compared to the use of whole animals in numbers sufficient for screening purposes. For the development of a system applicable to throughput screening formats, EB have been chosen as the model system for the determination of differentiation effects by biological agents. Since the goal of this study was to identify EB as a model system for differentiation studies by throughput methodologies, it was essential to develop a throughput-screening protocol using EB. For screening purposes, uniform-sized EB have been shown to have synchronous differentiation potentials (Ng et al. 2005). Therefore, a goal of this study was to determine uniformity of size of EB for use in throughput screen methodologies. Current techniques for EB studies include hanging drop and suspension methodologies (Dang et al. 2002; Gerecht-Nir et al. 2004). The suspension culture method gives rise to non uniformly sized cells (Brook and Gardner, 1997; Dang et al. 2002; Gerecht-Nir et al. 2004). The hanging drop method provides uniform sizes of EB; however, this technique is challenging to perform and not amenable to throughput screening strategies (Dang et al. 2002; Kurosawa et al. 2003). Current methods that ensure uniform sizes of EB in a screening format are not simple procedures and require the use of sophisticated flow sorting equipment (Fernández et al. 2005). Additionally, to dispense single EB into wells by currently available automated methods is not cost-effective. For these reasons, a simple, cost-effective method in which EB are generated of uniform size and singularity is herein described. Additionally, the EB are dispensable into micro-wells in a manner that enables throughput methodologies for screening purposes. The use of single EB in a multi-well format is ideal because theoretically, one can consider a single EB in a well as an equivalent to a developing embryo, or in other words, a single EB is a developing organism in a microwell. The method described here allows screening and testing of biochemical compounds, biological agents and infectious organisms as well as allowing for toxicity evaluation.
Materials and Methods
ES cells derived from 129/SvJ mice were maintained in culture on a layer of feeder cells consisting of mitomycin C-treated primary mouse embryonic fibroblasts (MEF) (Brook and Gardner, 1997). The ES cells were cultured in ES medium (DMEM supplemented with 15% serum (Hyclone, Logan, UT), 1000 units/ml LIF (ESGRO from Chemicon, Temecula, CA), 1mM sodium pyruvate, As a pre-differentiation step, feeder cells were removed from the ES cell culture. To accomplish feeder cell removal, ES cells were passaged three times onto 0.1% gelatin-coated tissue culture plates using the following protocol. The cells were trysinized (5 min) and trypsin was inactivated by addition of an equal volume of media followed by incubation in a humidified chamber (15 min, For differentiation, the ES cells on gelatin-coated tissue culture plates were washed with phosphate buffered saline (PBS), trypsinized, dissociated into single cells and suspended in differentiation medium (same formulation as ES medium except devoid of LIF). ES cells were counted using a hemocytometer and number of surviving cells was determined by trypan blue dye exclusion (> 99% cell viability). The final cell number was adjusted to 5 x 103 cells/ml. To achieve single EB with uniformity of size, 1,000 ES cells in a 200 μl volume were dispensed into sterile 96-well polyvinyl carbonate polymerase chain reaction (PCR), plates (Greiner Bio-One Inc, Longwood, FL), tapped gently and incubated ( For neuron differentiation, 4-dsEB were transferred to gelatin (0.1%)-coated 96-well tissue culture plates and exposed to 1 µM retinoic acid (Bain et al. 1995). After the 4th day of retinoic acid exposure, the wells received differentiation medium (formulation stated above). To verify EB differentiation, EB were dissociated and grown on coverslips coated with 0.1% gelatin in 6-well micro-plates, allowed to differentiate for 12 days and subjected to immunocytochemistry. To identify post-mitotic neurons, expression of a neuronal antigenic marker, microtubule associated neuron cytoskeletal protein (MAP2), was assessed by immunocytochemistry using antibody reactivity to MAP2 [a+b] (Sigma Chemical, Results were displayed as means ± standard error of the mean (SEM). Statistical analyses were by an unpaired, two-tailed Students t-test or by the Kruskal-Wallis test followed by Dunn’s multiple comparisons test when indicated.
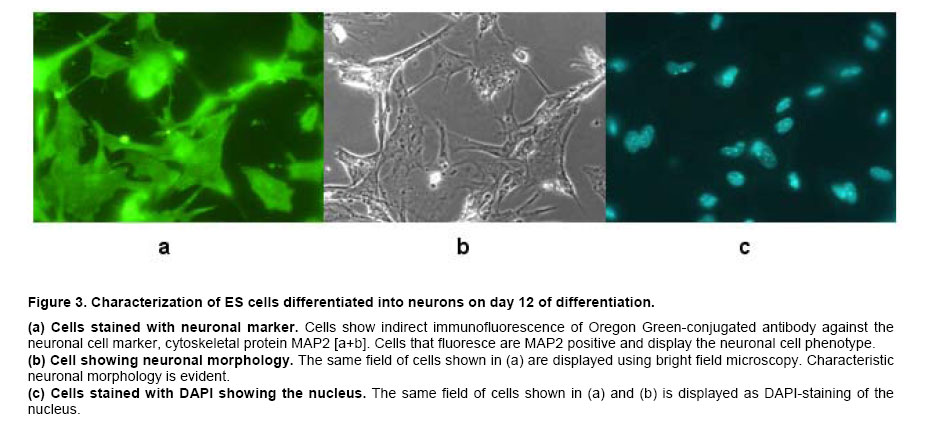
Results
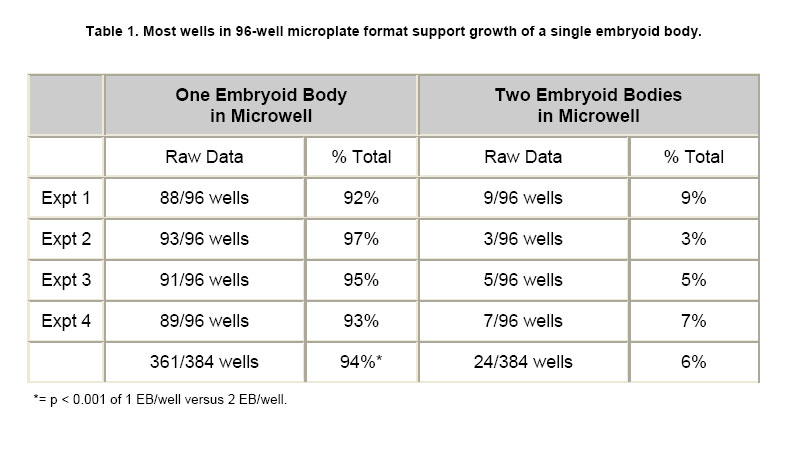
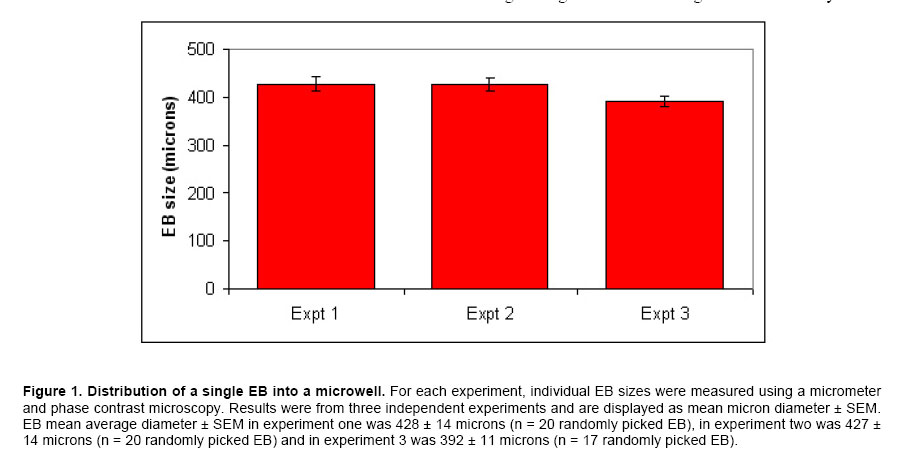
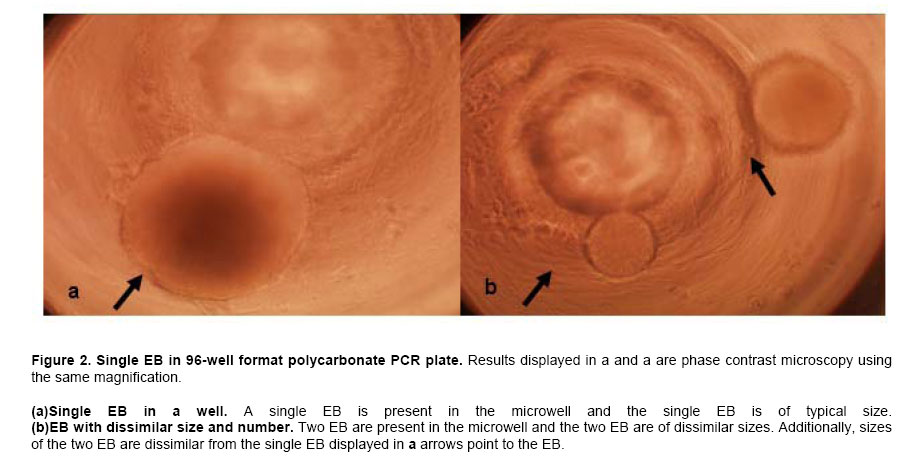
ES cells in 96-well PCR microplates were allowed to form EB. On the fourth day, the 4-dsEB were assessed as to number of EB per microwell. Table 1 shows results of four independent experiments to determine number of EB/well using the above-described method. All microwells contained either one or two EB per well with the predominant number of wells containing a single EB. Microwells containing more than two EB were never observed. In the four independent experiments, 92%, 97%, 95% and 93% (mean 94%) of the wells in 96-well multi-well plates contained a single EB. Two EB per well were observed 6% of the time (n = 4). To obtain a mean value for EB size from the 4-dsEB, single EB were randomly picked and EB diameters measured. Results from three independent experiments are displayed in Figure 1. The mean average diameter ± SEM of randomly picked EB in experiment one was 428 ± 14 microns (n = 20). In the second experiment, mean average diameter ± SEM of randomly picked EB was 427 ± 14 microns (n = 20) and in experiment 3, mean average diameter ± SEM of randomly picked EB was 392 ± 11 microns (n = 17). No statistically significant difference was identified among the three experiments. Mean average diameter of all three experiments (57 randomly picked EB) was 416 microns. Photomicrographs of EB in micro-well plates are displayed in Figure 2a single EB in micro-well format is displayed in Figure 2a and a well containing more than one EB is shown in Figure 2b. The EB were attached at either central or peripheral locations in the micro-well. As stated above, the most frequent observation was a single EB/well in this format. Rarely, two EB/well were evident. Every time two EB/well were present, the EB were either uniform or dissimilar in size by this method. Use of single EB/well in differentiation studies was demonstrated. It has been established in the literature that EB give rise to differentiated cardiac muscle cells (Doetschman et al. 1993; Muthuchamy et al. 1993; Kurosawa et al. 2003). Of importance, transfer of EB by the method reported here did not result in any mechanical injury-induced loss of EB as evidenced by the presence of intact cells of similar spherical morphology immediately after transfer as before manipulation. The EB in each well were periodically observed microscopically for generation of a unique characteristic, specifically of a beating cardiac muscle phenotype in the population of cells derived from the EB. A result was considered positive if an EB containing foci of a beating cardiac muscle was observed. A few EB showed foci of beating cells on day 3 of plating; however, by day 7, all EB in the wells were observed to have foci of beating cardiac muscle cells. In addition to cardiac-specific differentiation, use of the method described here allowed for the differentiation of EB into neuronal cells. The presence of neurite growth indicated that the EB in multi-well format had the ability to form neuronal cells. To verify the cells as neurons, indirect immunofluorescence for the presence of the MAP2 neuronal cell marker was performed on day 12-differentiated cells. Anti-MAP2 immunofluorescence positive cells with a typical neuronal morphology are shown in Figure 3a. Figure 3b displays the same field of cells as Figure 3abut with the use of bright field microscopy. Figure 3c shows results of the same field of cells (as displayed in Figure 3a and Figure 3b) but with DAPI-staining of the nucleus. The results indicate that on day 12 of differentiation, neuronal cells are present. EB formed from murine ES cells recapitulate many aspects of a developing embryo. Synchronous differentiation of EB is achieved by formation of uniformly sized EB (Ng et al. 2005). The method reported here is cost-effective and easy to perform. Additionally, a large number of EB can be generated using the 96-well PCR plate method followed by transfer of the EB to 96-well tissue culture plates for differentiation and screening purposes. On average, 94% of the wells have a single EB with mean average diameter of 415 microns. The EB formed by this method differentiate into different cell types with specific differentiation into beating cardiomyocytes and neurons being assessed in this study. The differentiation of ES cells into various lineages is an area of intense study because of the direct applicability of ES cells to the field of regenerative medicine (Smith, 2001; Ding and Schultz, 2004; Kumar et al. 2005). The in vitro differentiation capabilities of ES cells allow their use as a model system for the study of developmental potential as well as their use as a valuable reagent for stem cell therapeutic approaches. Several methodologies exist by which EB are formed and differentiated into different cell types. The typical method of EB formation is by removal of LIF and the feeder cell layer followed by growth in bacterial grade Petri dishes of EB in suspension. While this is a commonly used method, EB formation occurs but with great variability in EB shape and EB size (Doevendans et al. 2000; Dang et al. 2002). Several technologies for large-scale EB growth have been reported with examples being suspension cultures in bioreactors and encapsulation of ES cells in alginate microbeads or agarose capsules (Magyar et al. 2001; Dang et al. 2002; Gerecht-Nir et al. 2004; Konno et al. 2005). EB formation in the presence of feeder cells, specifically STO cells, and subsequent differentiation of these EB has been reported (Zhou et al. 2005). Currently, uniform-size single EB are usually generated by a hanging drop method in which the cells are allowed to aggregate in a hanging drop. This method is laborious and a paramount task is the acquisition of a sufficient number of EB for screening purposes (Kurosawa et al. 2003). The uniformity of size observed in EB formed by the hanging drop method has been suggested to be important for subsequent EB differentiation since endogenously produced LIF may exert an autocrine or paracrine function even though exogenous LIF is removed (Ansell and Hole, 2000). In the hanging drop method, an EB suspension containing approximately 300 cells in a small volume (10 μl) are placed onto the lid of a Petri plate. The Petri plate lid is placed onto its base with a swift action so as to not disperse the drop. The plate is kept in a humidified incubator (7.5% CO2 in air, 48 hrs). After two days, the cells have settled into the meniscus of the hanging drop and have formed smooth, spherical aggregates of uniform size. The EB are then pooled and used for differentiation studies. Since all the EB are present in the small droplet, an ever-present problem with the hanging drop method is that there is no easy way to subject individual EB to different biological agents. In the method described here, the EB settle to the bottom of the microwell in a multi-well plate. Slight tapping of the multi-well plate causes the EB to accumulate at the base of the well which then allows for single uniform-sized EB to be dispensed into the individual wells. Additionally, several polyvinyl carbonate PCR plates can be used to generate large numbers of single, uniform-sized EB which can then be transferred to multi-well tissue culture plates for subsequent differentiation and screening purposes. Herein we show that EB formed by this method are efficiently differentiated into cardiomyocytes and neurons. The formation of single, uniform-size EB in a multi-well format provides an efficient in vitro system to screen for effects of small molecules and bioagents and is an alternative to embryo and live animal studies. Using this in vitro system, it is possible to screen many compounds efficiently. Once a candidate molecule is identified, a thorough study can be performed to define how the molecule exerts its effect using throughput technology. To streamline throughput screening studies, ES cell lines containing a reporter construct under the control of a specific promoter in the form of a promoter-reporter cassette is of great interest. Reporters expressed only in lineages where the promoter is functional is an initial approach worth assessment. The differentiated cell type will be identifiable by analysis of reporter gene expression. Screening compounds may be added to EB and analysis of reporter expression would show whether the small molecule inhibits or augments differentiation (or other function) or has no effect on cell lineage differentiation. By creation of a panel of lineage-specific promoter-reporter ES cell lines, screening of EB from these cell lines has the potential to provide an excellent system for testing differentiation effects of small molecules. Currently, work is in progress to engineer ES cells lines useful for these types of aforementioned screening purposes. Formation of ES cells giving rise to uniform-sized, single EB is crucial for the success of this screening strategy. Information gleaned from this panel will be useful in determining whether a particular small molecule is toxic to the embryo or is a modulator of development of a particular cell lineage. Differentiation of ES cells into different cell lineages is of great importance since the potential exists for application of discovered technologies as therapeutics for regenerative medicine.
References
Note: Electronic Journal of Biotechnology is not responsible if on-line references cited on manuscripts are not available any more after the date of publication. © 2007 by Pontificia Universidad Católica de Valparaíso -- Chile The following images related to this document are available:Photo images[ej07031f3.jpg] [ej07031f2.jpg] [ej07031f1.jpg] [ej07031t1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}